



Effect of P62 on the migration and motility of human epidermal cell line HaCaT in high glucose microenvironment and its mechanism
-
摘要:
目的 研究高糖微环境下P62蛋白对人表皮细胞株HaCaT迁移和运动性的影响及其可能的分子机制,以探讨糖尿病足创面难愈合的机制。 方法 采用实验研究方法。取对数生长期HaCaT进行实验。取细胞,按随机数字表法(分组方法下同)分为正常对照组(培养液含终物质的量浓度5.5 mmol/L的葡萄糖)及高糖(培养液含终物质的量浓度30.0 mmol/L的葡萄糖)24 h组、高糖48 h组、高糖72 h组。正常对照组细胞行常规培养72 h,高糖72 h组细胞行高糖培养72 h,高糖48 h组细胞先常规培养24 h再高糖培养48 h,高糖24 h组细胞先常规培养48 h再高糖培养24 h后,采用蛋白质印迹法检测P62蛋白表达。取细胞,分为正常对照组、高糖组,分别同前培养48 h后,采用免疫荧光法检测P62蛋白表达(以绿色荧光表示)。取细胞,分为阴性对照小干扰RNA(siRNA)组、P62-siRNA-1组、P62-siRNA-2组、P62-siRNA-3组,并转染相应试剂,于转染后72 h,采用蛋白质印迹法检测P62蛋白表达。取细胞,分为正常糖+阴性对照siRNA组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组,并行相应处理,于转染后72 h,采用蛋白质印迹法检测P62蛋白表达;行划痕试验检测并计算划痕后24 h细胞迁移率(样本数为9);在活细胞工作站下,观察3 h内细胞运动范围并计算运动速度(正常糖+阴性对照siRNA组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组观察细胞数分别为76、75、80、79个)。取细胞,分为正常糖+磷酸盐缓冲液(PBS)组、高糖+PBS组、高糖+N-乙酰半胱氨酸(NAC)组,行相应处理后,于培养48 h,分别采用蛋白质印迹法及免疫荧光法检测P62蛋白表达。除划痕试验外,其余实验各组样本数均为3。对数据行单因素方差分析、LSD检验。 结果 与正常对照组比较,高糖24 h组、高糖48 h组及高糖72 h组细胞P62蛋白表达量均明显增加(P<0.01)。培养48 h,高糖组细胞中P62的绿色荧光强于正常对照组。转染后72 h,与阴性对照siRNA组比较,P62-siRNA-1组、P62-siRNA-2组和P62-siRNA-3组细胞P62蛋白表达量均明显减少(P<0.01)。转染后72 h,与正常糖+阴性对照siRNA组比较,正常糖+P62-siRNA组细胞P62蛋白表达量明显减少(P<0.01),高糖+阴性对照siRNA组细胞P62蛋白表达量明显增加(P<0.01);与高糖+阴性对照siRNA组比较,高糖+P62-siRNA组细胞P62蛋白表达量明显减少(P<0.01)。划痕后24 h,与正常糖+阴性对照siRNA组[(55±7)%]比较,正常糖+P62-siRNA组细胞迁移率明显升高[(72±14)%,P<0.01],高糖+阴性对照siRNA组细胞迁移率明显下降[(37±7)%,P<0.01];与高糖+阴性对照siRNA组比较,高糖+P62-siRNA组细胞迁移率明显升高[(54±10)%,P<0.01]。观察3 h内,高糖+阴性对照siRNA组细胞运动范围较正常糖+阴性对照siRNA组缩小,正常糖+P62-siRNA组细胞运动范围较正常糖+阴性对照siRNA组增大,高糖+P62-siRNA组细胞运动范围较高糖+阴性对照siRNA组增大。与正常糖+阴性对照siRNA组比较,正常糖+P62-siRNA组细胞运动速度明显增加(P<0.01),高糖+阴性对照siRNA组细胞运动速度明显下降(P<0.01);与高糖+阴性对照siRNA组比较,高糖+P62-siRNA组细胞运动速度明显增加(P<0.01)。培养48 h,与正常糖+PBS组比较,高糖+PBS组细胞P62蛋白表达量明显增加(P<0.01);与高糖+PBS组比较,高糖+NAC组细胞P62蛋白表达量明显减少(P<0.01)。培养48 h,高糖+PBS组细胞中P62的绿色荧光强于正常糖+PBS组,而高糖+NAC组细胞中P62的绿色荧光弱于高糖+PBS组。 结论 在HaCaT中,高糖微环境可促进P62蛋白表达;敲减P62蛋白可促进其迁移并增加运动性;高糖微环境下活性氧增加可能是P62表达增加的潜在机制。 Abstract:Objective To investigate the effect of P62 on the migration and motility of human epidermal cell line HaCaT in high glucose microenvironment and its possible molecular mechanism, so as to explore the mechanism of refractory diabetic foot wound healing. Methods The method of experimental research was used. HaCaT cells in logarithmic growth phase was taken for experiment. The cells were collected and divided into normal control group (culture solution containing glucose with final molarity of 5.5 mmol/L) and high glucose (culture solution containing glucose with final molarity of 30.0 mmol/L) 24 h group, high glucose 48 h group, and high glucose 72 h group according to the random number table (the same grouping method below). The cells in normal control group were routinely cultured for 72 h, cells in high glucose 72 h group were cultured with high glucose for 72 h, cells in high glucose 48 h group were routinely cultured for 24 h then cultured with high glucose for 48 h, cells in high glucose 24 h group were routinely cultured for 48 h then cultured with high glucose for 24 h. Then the protein expression of P62 was detected by Western blotting. The cells were collected and divided into normal control group and high glucose group. After being correspondingly cultured for 48 h as before, the protein expression of P62 was detected by immunofluorescence method (indicated as green fluorescence). The cells were collected and divided into negative control small interfering RNA (siRNA) group, P62-siRNA-1 group, P62-siRNA-2 group, and P62-siRNA-3 group, and transfected with the corresponding reagents. At post transfection hour (PTH) 72, the protein expression of P62 was detected by Western blotting. The cells were collected and divided into normal glucose+negative control siRNA group, normal glucose+P62-siRNA group, high glucose+negative control siRNA group, and high glucose+P62-siRNA group. After the corresponding treatment, the protein expression of P62 was detected by Western blotting at PTH 72 h, the cell migration rate was detected and calculated at 24 h after scratching by scratch test, with the number of samples being 9; and the range of cell movement was observed and the trajectory velocity was calculated within 3 h under the living cell workstation, with the number of samples being 76, 75, 80, and 79 in normal glucose+negative control siRNA group, normal glucose+P62-siRNA group, high glucose+negative control siRNA group, and high glucose+P62-siRNA group, respectively. The cells were collected and divided into normal glucose+phosphate buffered solution (PBS) group, high glucose+PBS group, and high glucose+N-acetylcysteine (NAC) group. After the corresponding treatment, the protein expression of P62 at 48 h of culture was detected by Western blotting and immunofluorescence method, respectively. Except for scratch test and cell motility experiment, the number of samples was all 3 in the rest experiments. Data were statistically analyzed with one-way analysis of variance and least significant difference test. Results Compared with the protein expression in normal control group, the protein expressions of P62 of cells in high glucose 24 h group, high glucose 48 h group, and high glucose 72 h group were significantly increased (P<0.01). At 48 h of culture, the green fluorescence of P62 of cells in high glucose group was stronger than that in normal control group. At PTH 72, compared with the protein expression in negative control siRNA group, the protein expressions of P62 of cells in P62-siRNA-1 group, P62-siRNA-2 group, and P62-siRNA-3 group were significantly decreased (P<0.01). At PTH 72, compared with the protein expression in normal glucose+negative control siRNA group, the protein expression of P62 of cells in normal glucose+P62-siRNA group was significantly decreased (P<0.01), while the protein expression of P62 of cells in high glucose+negative control siRNA group was significantly increased (P<0.01); compared with the protein expression in high glucose+negative control siRNA group, the protein expression of P62 of cells in high glucose+P62-siRNA group was significantly decreased (P<0.01). At 24 h after scratching, compared with (55±7)% in normal glucose+negative control siRNA group, the cell migration rate in normal glucose+P62-siRNA group was significantly increased ((72±14)%, P<0.01), while the cell migration rate in high glucose+negative control siRNA group was significantly decreased ((37±7)%, P<0.01); compared with that in high glucose+negative control siRNA group, the cell migration rate in high glucose+P62-siRNA group was significantly increased ((54±10)%, P<0.01). Within 3 h of observation, the cell movement range in high glucose+negative control siRNA group was smaller than that in normal glucose+negative control siRNA group, while the cell movement range in normal glucose+P62-siRNA group was larger than that in normal glucose+negative control siRNA group, and the cell movement range in high glucose+P62-siRNA group was larger than that in high glucose+negative control siRNA group. Compared with that in normal glucose+negative control siRNA group, the cell trajectory speed in normal glucose+P62-siRNA group was significantly increased (P<0.01), while the cell trajectory speed in high glucose+negative control siRNA group was significantly decreased (P<0.01); compared with that in high glucose+negative control siRNA group, the cell trajectory speed in high glucose+P62-siRNA group was significantly increased (P<0.01). At 48 h of culture, compared with that in normal glucose+PBS group, the protein expression of P62 of cells in high glucose+PBS group was significantly increased (P<0.01); compared with that in high glucose+PBS group, the protein expression of P62 of cells in high glucose+NAC group was significantly decreased (P<0.01). At 48 h of culture, the green fluorescence of P62 of cells in high glucose+PBS group was stronger than that in normal glucose+PBS group, while the green fluorescence of P62 of cells in high glucose+NAC group was weaker than that in high glucose+PBS group. Conclusions In HaCaT cells, high glucose microenvironment can promote the protein expression of P62; knockdown of P62 protein can promote the migration and increase the mobility of HaCaT cells; and the increase of reactive oxygen species in high glucose microenvironment may be the underlying mechanism for the increase of P62 expression. -
Key words:
- Diabetic foot /
- Reactive oxygen species /
- Cell migration assays /
- P62 /
- High glucose /
- Wound repair
-
(1)观察到高糖微环境下人表皮细胞株HaCaT中P62蛋白表达增加,敲减P62蛋白可促进HaCaT的迁移并增加其运动性。
(2)初步证明活性氧可能是高糖微环境下HaCaT中P62表达增加的潜在上游机制。
糖尿病是一组以高血糖为主要标志的、因胰岛素绝对或相对分泌不足以及靶组织细胞对胰岛素敏感性下降引起的代谢紊乱综合征[1]。糖尿病足是导致我国糖尿病患者残疾、死亡的严重慢性并发症之一,发病率高,治疗困难且花费巨大,因此,研究并揭示糖尿病创面愈合不良的潜在分子机制是一个至关重要的公共卫生问题[2, 3]。
自噬是一种通过自噬小体将不需要或老化受损的细胞器和蛋白质靶向到溶酶体中进行降解的自我更新机制[4, 5, 6]。P62蛋白是一种多结构域蛋白,是细胞自噬过程中选择吞噬物的衔接分子和底物[7]。P62通过微管相关蛋白1轻链3(LC3)相互作用区与自噬体膜中的LC3结合,以及通过泛素相关结构域结合降解的多泛素化蛋白和蛋白多聚体[8]。P62在被发现之初是一种与非典型蛋白激酶C相互作用的蛋白,在氧化应激反应中发挥重要作用,同时对细胞增殖、生长和存活具有调节作用。近年来研究表明,P62除了作为自噬衔接分子发挥作用外,还参与肺癌、乳腺癌、黑色素瘤、皮肤鳞癌等肿瘤的发生和进展[9, 10, 11]。此外,P62蛋白异常表达还与神经退行性病变、感染性疾病、遗传性疾病等的发生发展密切相关[12, 13]。本课题组前期研究结果表明,高糖条件下人KC中的自噬受到明显抑制,P62表达增加[14]。然而P62蛋白在皮肤KC中的作用鲜有报道。本实验研究通过构建小干扰RNA(siRNA)敲减P62蛋白,检测高糖微环境下P62对皮肤KC迁移及运动性的影响,以探讨其在糖尿病创面愈合中的作用及潜在机制。
1. 材料与方法
1.1 细胞及主要试剂与仪器来源
人永生化表皮细胞株HaCaT购自中国科学院上海细胞生物学研究所。胎牛血清购自美国Gibco公司,改良型RPMI 1640培养液、青/链霉素购自美国HyClone公司,P62-siRNA-1、P62-siRNA-2、P62-siRNA-3由上海吉玛制药技术有限公司构建,兔抗小鼠P62多克隆抗体购自美国CST公司,辣根过氧化物酶(HRP)标记的兔抗小鼠GAPDH多克隆抗体、HRP标记的山羊抗兔IgG多克隆抗体、异硫氰酸荧光素(FITC)标记的山羊抗兔IgG多克隆抗体购自美国Proteintech公司,N-乙酰半胱氨酸(NAC)、4',6-二脒基-2-苯基吲哚(DAPI)购自美国Sigma公司。LSM510 Meta型活细胞工作站购自德国Zeiss公司,ChemiDoc XRS型凝胶成像仪购自美国Bio-Rad公司,TCS SP5型激光扫描共聚焦显微镜购自德国Leica公司,DP80型倒置光学显微镜购自日本Olympus公司。
1.2 细胞培养
将含100 U/mL青霉素/链霉素、体积分数为10%胎牛血清的改良型RPMI 1640培养液(以下简称完全培养液,含终物质的量浓度为5.5 mmol/L的葡萄糖)加入底面积25 cm2的培养瓶后,将HaCaT接种于该培养瓶中,于37 ℃、含体积分数5%二氧化碳的细胞培养箱中培养(以下简称常规培养),隔1 d换液1次。当细胞生长达80%~90%融合时传代。取对数生长期细胞用于后续实验。
1.3 高糖微环境下HaCaT中P62蛋白表达变化
1.3.1 蛋白质印迹法检测
取细胞,按照浓度1×106个/mL接种于6孔板中,每孔2 mL。采用随机数字表法(分组方法下同)将细胞分为正常对照组、高糖24 h组、高糖48 h组及高糖72 h组,每组1孔。对正常对照组细胞行常规培养72 h,高糖72 h组细胞行高糖培养(完全培养液中含终物质的量浓度为30.0 mmol/L的葡萄糖)72 h,高糖24 h组细胞先行常规培养48 h再行高糖培养24 h,高糖48 h组细胞先行常规培养24 h再行高糖培养48 h。培养72 h后,采用蛋白质印迹法检测细胞P62蛋白表达。加入的一抗为兔抗小鼠P62多克隆抗体(稀释比为1∶1 000)、HRP标记的兔抗小鼠GAPDH多克隆抗体(稀释比为1∶5 000),二抗为HRP标记的山羊抗兔IgG多克隆抗体(稀释比为1∶5 000)。以GAPDH为内参照,计算目的蛋白与内参照蛋白灰度值的比值。该实验重复3次。
1.3.2 免疫荧光法检测
将细胞以每孔5×104个的密度接种于24孔板的细胞爬片上。常规培养24 h后,将细胞分为正常对照组、高糖组,每组1孔,分别同1.3.1行常规培养和高糖培养。培养48 h,取细胞以40 g/L多聚甲醛固定并采用免疫荧光法检测P62蛋白表达,其中一抗为兔抗小鼠P62多克隆抗体(稀释比为1∶100)、二抗为FITC标记的山羊抗兔IgG多克隆抗体(稀释比为1∶200),采用DAPI进行细胞核染色(阳性为蓝色)。于630倍激光扫描共聚焦显微镜下观察并每孔选择3个视野获取图像,P62阳性染色为绿色。该实验重复3次,选择最有代表性的1次结果进行展示。
1.4 高糖微环境下HaCaT中P62蛋白敲减效果的验证
1.4.1 蛋白质印迹法筛选对P62蛋白敲减效果最佳的siRNA
将细胞以每孔1×105个的密度接种于6孔板中常规培养。当细胞生长达60%~80%融合时,分组并采用脂质体2000进行siRNA转染,阴性对照siRNA组(正义链:5'-UUCUCCGAACGUGUCACUUTT-3',反义链:5'-ACGUGACACGUUCGGAGAATT-3')、P62-siRNA-1组(正义链:5'-CCAGACUACGACUUGUGUATT-3',反义链:5'-UACACAAGUCGUAGUCUGGTT-3')、P62-siRNA-2组(正义链:5'-CCUACGUGAAGGAUGACAUTT-3',反义链:5'-AUGUCAUCCUUCACGUAGGTT-3')、P62-siRNA-3组(正义链:5'-GGAACAGAUGGAGUCGGAUTT-3',反义链:5'-AUCCGACUCCAUCUGUUCCTT-3'),每组1孔。转染后72 h,同1.3.1采用蛋白质印迹法检测P62蛋白表达,明确siRNA的敲减效果,并选择效果最佳者用于后续实验。该实验重复3次。
1.4.2 蛋白质印迹法检测高糖微环境下siRNA对P62蛋白的敲减效果
取细胞按1×106个/mL的浓度接种于6孔板中,每孔2 mL。常规培养24 h后,将细胞分为正常糖+阴性对照siRNA组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组,每组1孔。正常糖+阴性对照siRNA组和正常糖+P62-siRNA组细胞分别转染阴性对照siRNA和P62-siRNA;高糖+阴性对照siRNA组、高糖+P62-siRNA组细胞分别转染阴性对照siRNA和P62-siRNA 24 h后行高糖培养48 h,其中siRNA转染方法同1.4.1,高糖培养方法同1.3.1。转染后72 h,同1.3.1采用蛋白质印迹法检测细胞中P62蛋白表达。该实验重复3次。
1.5 高糖微环境下P62对HaCaT迁移的影响
采用划痕试验[15]检测。将细胞以每孔4×105个的密度接种于6孔板中,细胞分组及处理同1.4.2,每组3孔。采用规格为100 μL的移液器枪头在培养孔正中垂直于培养孔划一竖痕,并使划痕细胞完全脱落。将细胞置于100倍倒置光学显微镜下拍照。采用ImageJ 1.51图像分析软件(美国国立卫生研究院)测量划痕后0(即刻)、24 h剩余划痕面积,计算划痕后24 h细胞迁移率。细胞迁移率=(划痕后0 h划痕面积-划痕后24 h剩余划痕面积)÷划痕后0 h面积×100%。该实验重复3次,结果取均值。
1.6 高糖微环境下P62对HaCaT运动性的影响
将细胞以每孔5×103个的密度接种于24孔板,细胞分组及处理同1.4.2,每组1孔。常规培养24 h后,于活细胞工作站倒置相差显微镜100倍镜下观察记录3 h内细胞运动情况,每5分钟记录1次图像,观察3 h内单个细胞运动情况,正常糖+阴性对照siRNA组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组观察细胞数分别为76、75、80、79个[10]。然后采用ImageJ 1.5.1图像分析软件分析细胞运动范围(以运动轨迹表示)和运动速度。细胞运动速度=细胞运动轨迹的总长度÷细胞运动时间。该实验重复3次。
1.7 高糖微环境下活性氧对HaCaT中P62蛋白表达的影响
1.7.1 蛋白质印迹法检测
将细胞以每孔1×106个的密度接种于6孔板。常规培养24 h后,将细胞分为正常糖+PBS组、高糖+PBS组、高糖+NAC组,每组1孔。高糖+NAC组细胞中加入NAC并调整其终物质的量浓度为5 mmol/L,正常糖+PBS组、高糖+PBS组细胞加入相同体积的PBS,高糖培养方法同1.3.1。培养48 h,同1.3.1采用蛋白质印迹法检测P62蛋白表达。该实验重复3次。
1.7.2 免疫荧光法检测
将细胞以每孔5×104个的密度接种于24孔板中的细胞爬片上。细胞分组及处理同1.7.1,每组1孔。各组细胞常规培养48 h后同1.3.2采用免疫荧光法检测P62蛋白表达。该实验重复3次。
1.8 统计学处理
采用SPSS 13.0统计软件进行数据分析。计量资料数据均符合正态分布,以
2. 结果
2.1 高糖微环境下HaCaT中P62蛋白表达变化
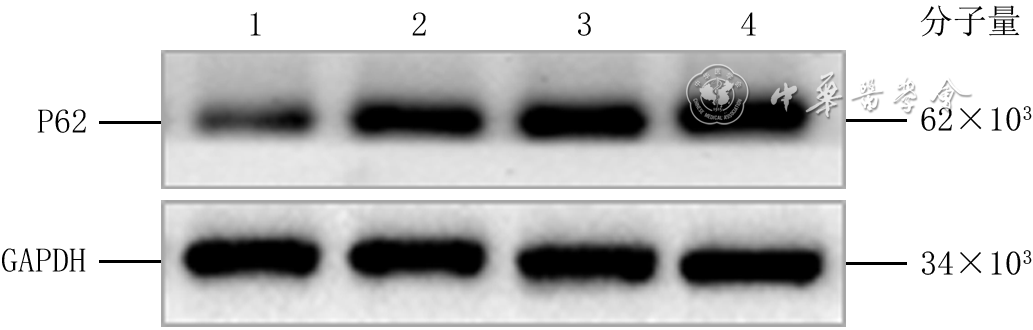
正常对照组、高糖24 h组、高糖48 h组和高糖72 h组细胞P62蛋白表达量分别为0.32±0.09、0.83±0.14、0.70±0.13、0.81±0.16,组间总体比较,差异有统计学意义(F=14.81,P<0.001)。与正常对照组比较,高糖24 h组、高糖48 h组及高糖72 h组细胞P62蛋白表达量均明显增加(P值分别为<0.001、0.001、<0.001);高糖24 h组与高糖48 h组、高糖24 h组与高糖72 h组、高糖48 h组与高糖72 h组细胞P62蛋白表达量均相近(P值分别为0.078、0.821、0.120)。见图1。
 1 蛋白质印迹法检测正常对照组和高糖培养3个时间点组HaCaT中P62蛋白表达注:GAPDH为3-磷酸甘油醛脱氢酶;条带上方1、2、3、4分别指示正常对照组、高糖24 h组、高糖48 h组、高糖72 h组
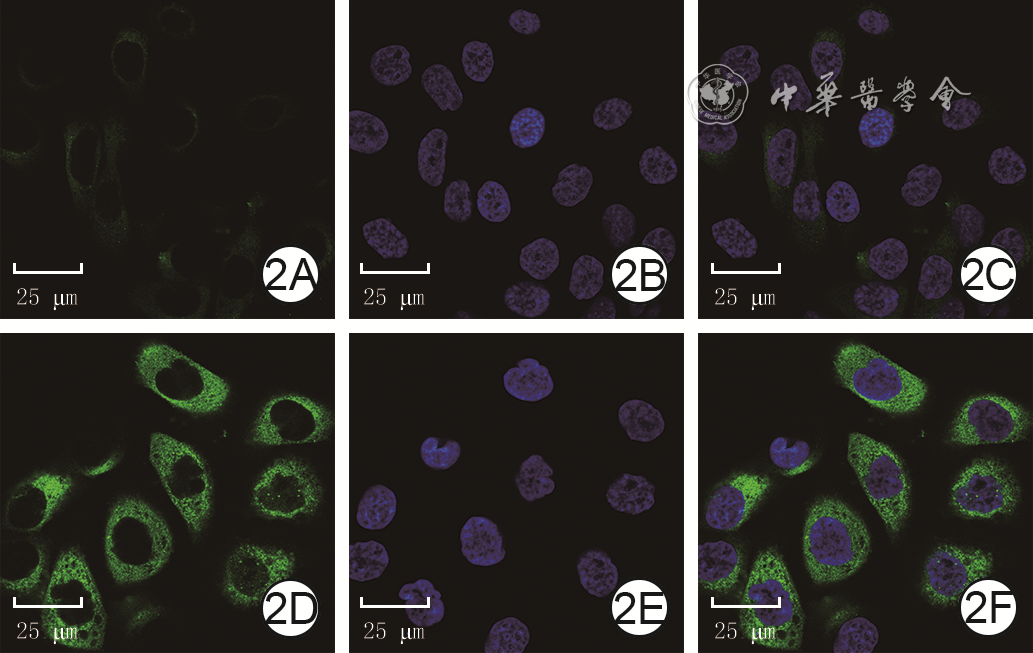
1 蛋白质印迹法检测正常对照组和高糖培养3个时间点组HaCaT中P62蛋白表达注:GAPDH为3-磷酸甘油醛脱氢酶;条带上方1、2、3、4分别指示正常对照组、高糖24 h组、高糖48 h组、高糖72 h组培养48 h,正常对照组细胞中P62的绿色荧光较弱,高糖组细胞中P62的绿色荧光强于正常对照组。见图2。
 2 正常对照组和高糖组HaCaT培养48 h的P62蛋白表达 异硫氰酸荧光素-4',6-二脒基-2-苯基吲哚×630。2A、2B、2C.分别为正常对照组P62染色、细胞核染色、P62与细胞核染色重叠图片,细胞核完整,P62蛋白表达较少,主要分布于细胞质;2D、2E、2F.分别为高糖组P62染色、细胞核染色、P62与细胞核染色重叠图片,细胞核完整,P62蛋白表达较图2A多注:P62阳性染色为绿色,细胞核阳性染色为蓝色
2 正常对照组和高糖组HaCaT培养48 h的P62蛋白表达 异硫氰酸荧光素-4',6-二脒基-2-苯基吲哚×630。2A、2B、2C.分别为正常对照组P62染色、细胞核染色、P62与细胞核染色重叠图片,细胞核完整,P62蛋白表达较少,主要分布于细胞质;2D、2E、2F.分别为高糖组P62染色、细胞核染色、P62与细胞核染色重叠图片,细胞核完整,P62蛋白表达较图2A多注:P62阳性染色为绿色,细胞核阳性染色为蓝色2.2 siRNA敲减HaCaT中P62蛋白的表达
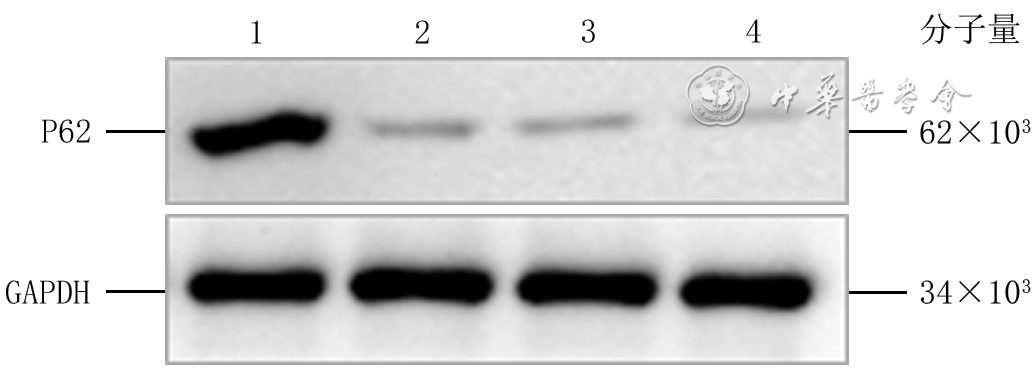
转染后72 h,阴性对照siRNA组、P62-siRNA-1组、P62-siRNA-2组和P62-siRNA-3组细胞P62蛋白表达量分别为0.970±0.189、0.217±0.192、0.099±0.024、0.066±0.030,组间总体比较,差异有统计学意义(F=39.16,P<0.001)。与阴性对照siRNA组比较,P62-siRNA-1组、P62-siRNA-2组和P62-siRNA-3组细胞P62蛋白表达量均明显减少(P<0.001);P62-siRNA-1组与P62-siRNA-2组、P62-siRNA-1组与P62-siRNA-3组、P62-siRNA-2组与P62-siRNA-3组细胞P62蛋白表达量均相近(P值分别为0.243、0.142、0.738)。见图3。
 3 蛋白质印迹法检测转染siRNA后4组HaCaT中P62蛋白表达注:GAPDH为3-磷酸甘油醛脱氢酶,siRNA为小干扰RNA;条带上方1、2、3、4分别指示阴性对照siRNA组、P62-siRNA-1组、P62-siRNA-2组、P62-siRNA-3组
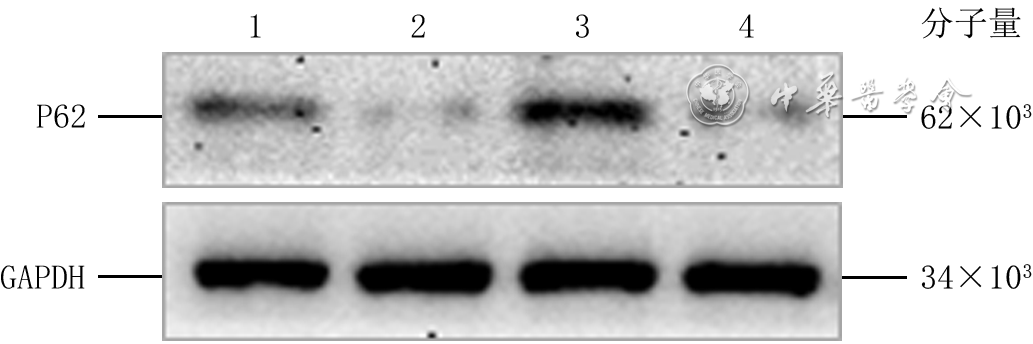
3 蛋白质印迹法检测转染siRNA后4组HaCaT中P62蛋白表达注:GAPDH为3-磷酸甘油醛脱氢酶,siRNA为小干扰RNA;条带上方1、2、3、4分别指示阴性对照siRNA组、P62-siRNA-1组、P62-siRNA-2组、P62-siRNA-3组选择目的片段P62-siRNA-3作为进一步研究的siRNA片段。转染后72 h,正常糖+阴性对照siRNA组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组和高糖+P62-siRNA组细胞P62蛋白表达量分别为0.66±0.27、0.30±0.18、1.14±0.20、0.45±0.17,组间总体比较,差异有统计学意义(F=18.56,P<0.001)。与正常糖+阴性对照siRNA组比较,正常糖+P62-siRNA组细胞P62蛋白表达量明显减少(P=0.006),高糖+阴性对照siRNA组细胞P62蛋白表达量明显增加(P=0.001);与高糖+阴性对照siRNA组比较,高糖+P62-siRNA组细胞P62蛋白表达量明显减少(P<0.001)。见图4。
 4 蛋白质印迹法检测高糖微环境下siRNA对4组HaCaT转染后72 h的P62蛋白表达的作用注:GAPDH为3-磷酸甘油醛脱氢酶,siRNA为小干扰RNA;条带上方1、2、3、4分别指示正常糖+阴性对照siRNA组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组
4 蛋白质印迹法检测高糖微环境下siRNA对4组HaCaT转染后72 h的P62蛋白表达的作用注:GAPDH为3-磷酸甘油醛脱氢酶,siRNA为小干扰RNA;条带上方1、2、3、4分别指示正常糖+阴性对照siRNA组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组2.3 高糖微环境下P62对HaCaT迁移的影响
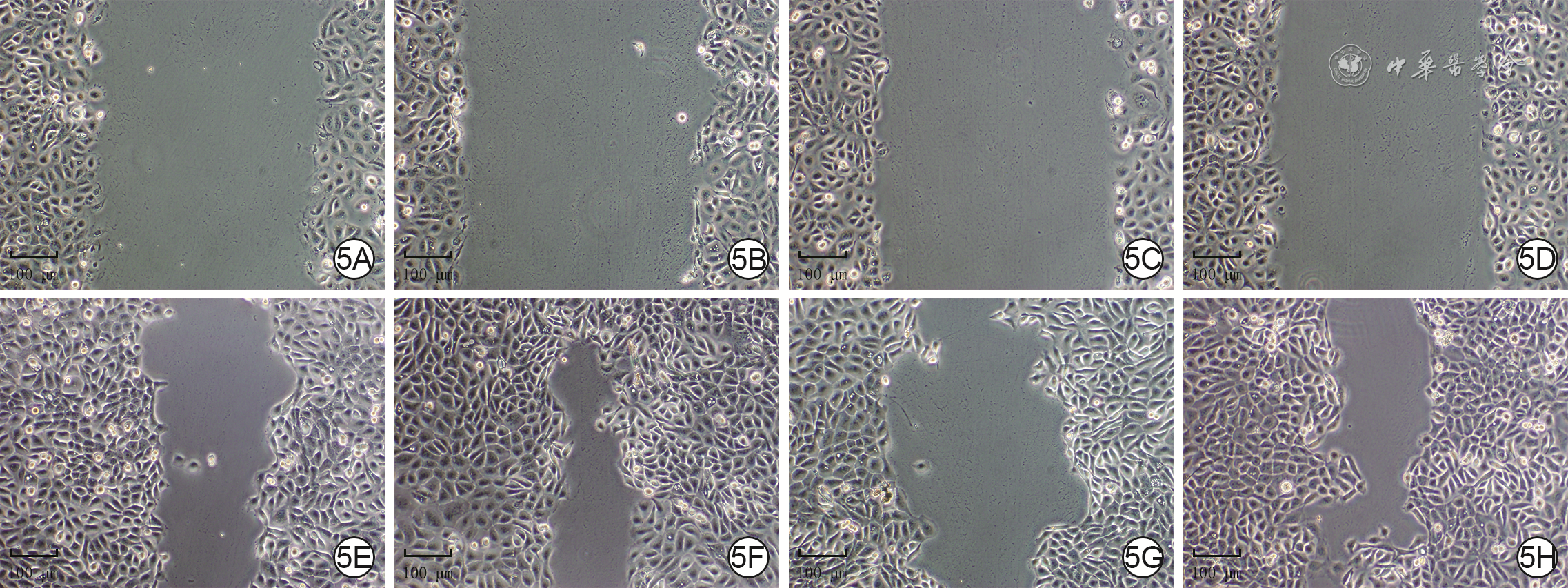
划痕后24 h,正常糖+阴性对照siRNA组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组细胞迁移率分别为(55±7)%、(72±14)%、(37±7)%、(54±10)%,组间总体比较,差异有统计学意义(F=18.20,P<0.001)。与正常糖+阴性对照siRNA组比较,正常糖+P62-siRNA组细胞迁移率明显升高(P=0.001),高糖+阴性对照siRNA组细胞迁移率明显下降(P<0.001);与高糖+阴性对照siRNA组比较,高糖+P62-siRNA组细胞迁移率明显升高(P=0.001)。见图5。
 5 4组HaCaT处理完成后划痕后各时间点细胞划痕面积情况 倒置光学显微镜×100。5A、5B、5C、5D.分别为正常糖+阴性对照小干扰RNA(siRNA)组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组划痕后0 h(即刻),划痕情况相近;5E、5F、5G、5H.分别为正常糖+阴性对照siRNA组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组划痕后24 h,图5F划痕面积明显小于图5E,图5G划痕面积明显大于图5E,图5H划痕面积明显小于图5G
5 4组HaCaT处理完成后划痕后各时间点细胞划痕面积情况 倒置光学显微镜×100。5A、5B、5C、5D.分别为正常糖+阴性对照小干扰RNA(siRNA)组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组划痕后0 h(即刻),划痕情况相近;5E、5F、5G、5H.分别为正常糖+阴性对照siRNA组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组划痕后24 h,图5F划痕面积明显小于图5E,图5G划痕面积明显大于图5E,图5H划痕面积明显小于图5G2.4 高糖微环境下P62对HaCaT运动性的影响
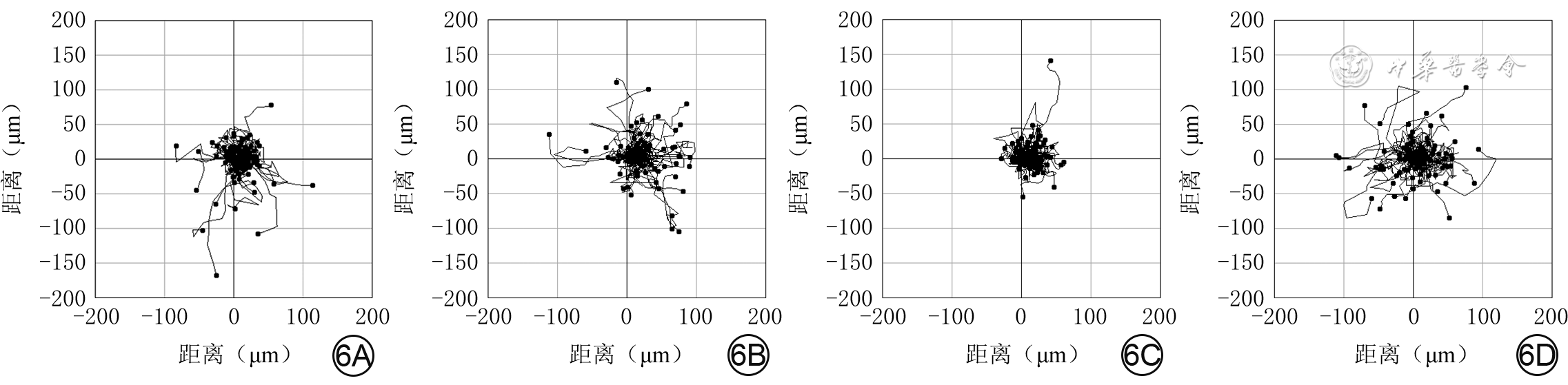
观察3 h内,高糖+阴性对照siRNA组细胞运动范围较正常糖+阴性对照siRNA组缩小,正常糖+P62-siRNA组细胞运动范围较正常糖+阴性对照siRNA组增大,高糖+P62-siRNA组细胞运动范围较高糖+阴性对照siRNA组增大。见图6。
 6 活细胞工作站观察4组HaCaT 3 h内的运动范围。6A、6B、6C、6D.分别为正常糖+阴性对照siRNA组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组,其中图6B、6D运动范围分别较图6A、6C明显增大注:siRNA为小干扰RNA;细胞运动起点均为坐标(0,0),运动终点为位于4个象限中的圆点,连接两者之间的曲线为细胞运动轨迹
6 活细胞工作站观察4组HaCaT 3 h内的运动范围。6A、6B、6C、6D.分别为正常糖+阴性对照siRNA组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组,其中图6B、6D运动范围分别较图6A、6C明显增大注:siRNA为小干扰RNA;细胞运动起点均为坐标(0,0),运动终点为位于4个象限中的圆点,连接两者之间的曲线为细胞运动轨迹观察3 h内,正常糖+阴性对照siRNA组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组细胞运动速度分别为(0.49±0.20)、(0.65±0.27)、(0.41±0.11)、(0.63±0.31)μm/min,组间总体比较,差异有统计学意义(F=39.04,P<0.001)。与正常糖+阴性对照siRNA组比较,正常糖+P62-siRNA组细胞运动速度明显增加(P<0.001),高糖+阴性对照siRNA组细胞运动速度明显下降(P=0.001);与高糖+阴性对照siRNA组比较,高糖+P62-siRNA组细胞运动速度明显增加(P<0.001)。
2.5 高糖微环境下活性氧对HaCaT中P62蛋白表达的影响
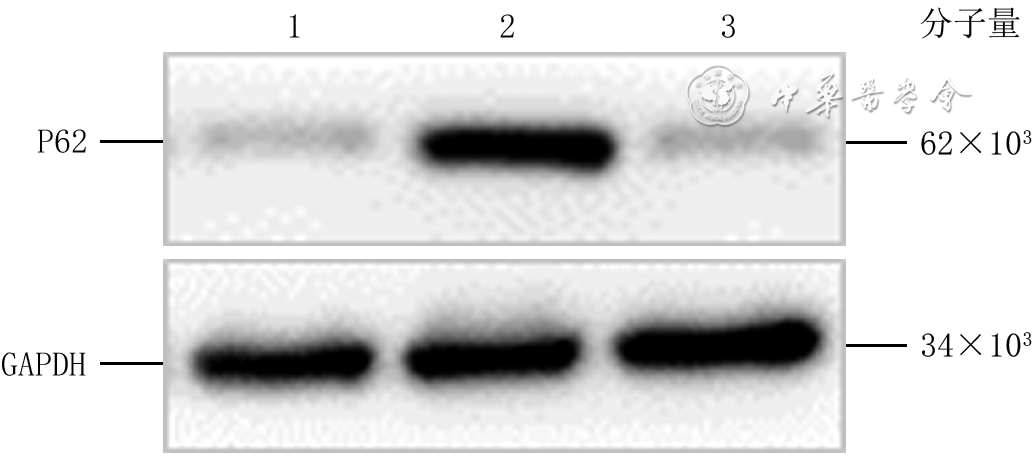
培养48 h,正常糖+PBS组、高糖+PBS组、高糖+NAC组细胞P62蛋白表达量分别为0.49±0.05、1.07±0.05、0.53±0.06,组间总体比较,差异有统计学意义(F=13.23,P=0.001)。与正常糖+PBS组比较,高糖+PBS组细胞P62蛋白表达量明显增加(P=0.001);与高糖+PBS组比较,高糖+NAC组细胞P62蛋白表达量明显减少(P=0.001)。见图7。
 7 蛋白质印迹法检测高糖微环境下NAC对3组HaCaT培养48 h的P62蛋白表达的影响注:GAPDH为3-磷酸甘油醛脱氢酶,PBS为磷酸缓冲盐溶液,NAC为N-乙酰半胱氨酸;条带上方1、2、3分别指示正常糖+PBS组、高糖+PBS组、高糖+NAC组
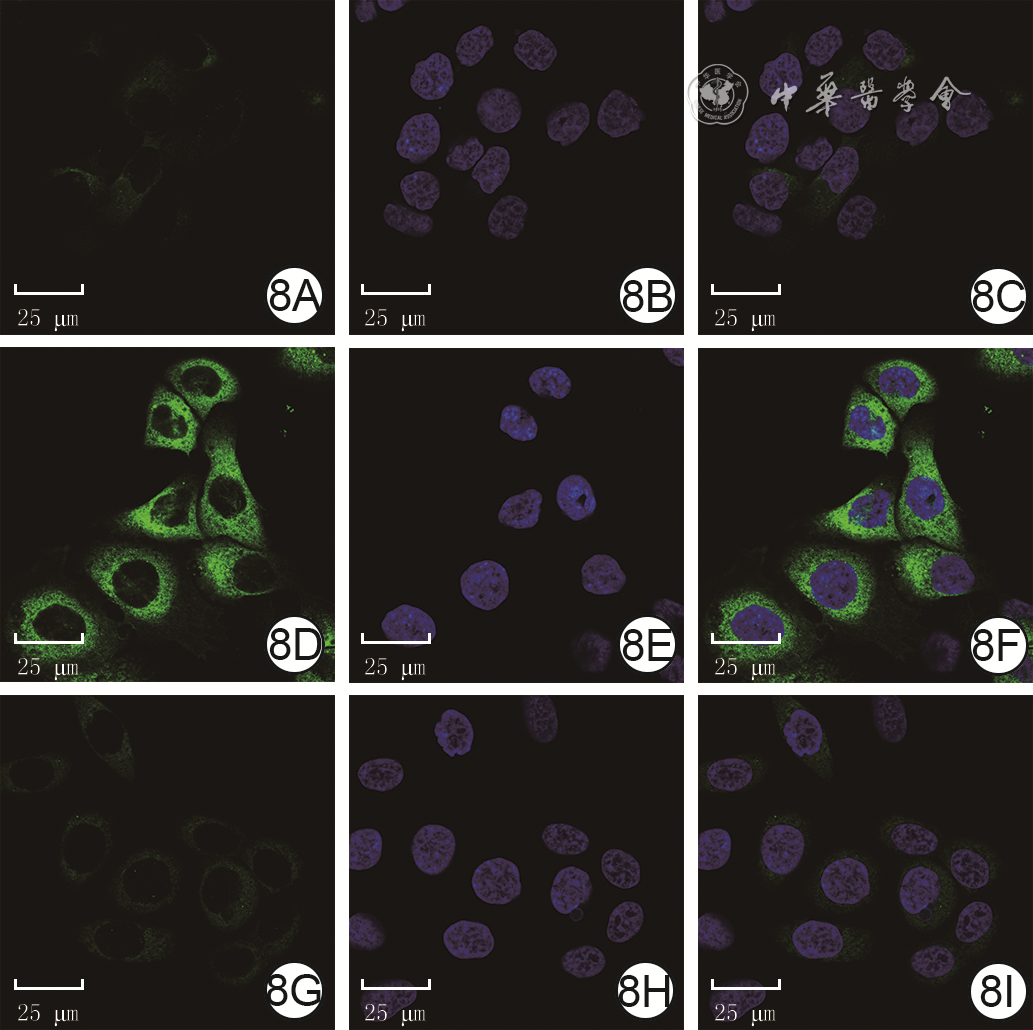
7 蛋白质印迹法检测高糖微环境下NAC对3组HaCaT培养48 h的P62蛋白表达的影响注:GAPDH为3-磷酸甘油醛脱氢酶,PBS为磷酸缓冲盐溶液,NAC为N-乙酰半胱氨酸;条带上方1、2、3分别指示正常糖+PBS组、高糖+PBS组、高糖+NAC组培养48 h,正常糖+PBS组细胞中P62的绿色荧光较弱,高糖+PBS组细胞中P62的绿色荧光强于正常糖+PBS组,而高糖+NAC组细胞中P62的绿色荧光弱于高糖+PBS组。见图8。
 8 3组HaCaT培养48 h的P62蛋白表达 异硫氰酸荧光素-4',6-二脒基-2-苯基吲哚×630。8A、8B、8C.分别为正常糖+PBS组P62染色、细胞核染色、P62与细胞核染色重叠图片,细胞核完整,P62蛋白表达较少,主要分布于细胞质;8D、8E、8F.分别为高糖+PBS组P62染色、细胞核染色、P62与细胞核染色重叠图片,细胞核完整,P62蛋白表达较图8A多;8G、8H、8I.分别为高糖+NAC组P62染色、细胞核染色、P62与细胞核染色重叠图片,细胞核完整,P62蛋白表达较图8D少注:P62阳性染色为绿色,细胞核阳性染色为蓝色;PBS为磷酸缓冲盐溶液,NAC为N-乙酰半胱氨酸
8 3组HaCaT培养48 h的P62蛋白表达 异硫氰酸荧光素-4',6-二脒基-2-苯基吲哚×630。8A、8B、8C.分别为正常糖+PBS组P62染色、细胞核染色、P62与细胞核染色重叠图片,细胞核完整,P62蛋白表达较少,主要分布于细胞质;8D、8E、8F.分别为高糖+PBS组P62染色、细胞核染色、P62与细胞核染色重叠图片,细胞核完整,P62蛋白表达较图8A多;8G、8H、8I.分别为高糖+NAC组P62染色、细胞核染色、P62与细胞核染色重叠图片,细胞核完整,P62蛋白表达较图8D少注:P62阳性染色为绿色,细胞核阳性染色为蓝色;PBS为磷酸缓冲盐溶液,NAC为N-乙酰半胱氨酸3. 讨论
创面愈合是一个多环节的、动态的、相互协调的生物学过程,涉及炎症反应、再上皮化和组织重塑。再上皮化是创面愈合过程中的重要环节,受KC迁移和增殖的调控[16, 17, 18, 19]。糖尿病状态下,持续的高糖环境导致病理性的皮肤组织学和细胞生物学行为的改变,例如蛋白质的氨基与糖的醛基之间的非酶促糖基化反应明显增加,形成晚期糖基化终末产物,氧化应激反应增强,毒害作用不断放大[20, 21]。皮肤KC功能障碍是导致糖尿病创面愈合不良的潜在关键因素,而糖尿病高糖微环境是导致皮肤KC迁移受损的重要因素之一[22]。虽然已有一系列关于糖尿病创面愈合的研究,但糖尿病创面修复的技术手段仍较局限,导致KC迁移受损的潜在分子机制仍需进一步深入探讨。
P62是一种细胞内应激诱导蛋白,与多种信号蛋白相互作用,可调节多种细胞功能。越来越多的研究证实,P62不仅参与自噬过程,还具有许多非自噬功能。例如,P62在肿瘤发生发展中的重要作用,有望成为肿瘤治疗的靶点[23]。P62具有多个结构域,包括氨基端的Phox1-Bem1p结构域[24]、ZZ型锌指结构域[24]、TNF受体相关因子6结合结构域[25]、LIR结构域[25]、Kelch样ECH相关蛋白1相互作用区和羧基端UBA结构域[26]。因此,P62可发挥多种多样的功能。本研究显示,高糖微环境下HaCaT中P62蛋白表达的显著增加。
细胞迁移一般指单个细胞、细胞片层和细胞群集从某一个位置到另一个位置的实际运动;细胞运动性则侧重于细胞运动的协调性,其目的性相对较差[27, 28]。本研究采用细胞划痕试验及单个细胞运动性实验2种方法,分别从细胞片层和单个细胞2个不同的维度检测了高糖微环境下KC迁移和运动性的改变;采用siRNA转染技术特异性敲减HaCaT中P62蛋白的表达。进一步实验结果表明,正常糖+P62-siRNA组及高糖+P62-siRNA组分别较正常糖+阴性对照siRNA组及高糖+阴性对照siRNA组中KC的细胞迁移率增加、细胞运动范围增大、运动速度增加,这提示敲减P62蛋白可改善皮肤KC的迁移和运动性。
高糖微环境可引起线粒体新陈代谢紊乱,从而产生大量活性氧。活性氧是糖尿病创面愈合的重要调节因子,低剂量的活性氧有利于抵抗外界损伤,而过量的活性氧则导致氧化还原失衡,是造成糖尿病创面不愈合的主要原因[29, 30]。活性氧可整合核因子κB、MAPK、磷脂酰肌醇3激酶/蛋白激酶B、核转录因子红系2相关因子2等多条信号转导通路以调控靶蛋白的表达,从而调节细胞的增殖、迁移、凋亡与自噬等生物学行为[31, 32]。研究显示,在人绒毛膜癌细胞株JEG-3中活性氧可通过c-Jun氨基末端蛋白激酶信号通路促进P62的表达,进而调节细胞凋亡[33]。由此本研究团队推测,在KC中活性氧可能对P62的表达具有调控作用。NAC是一种有效的活性氧清除剂[34],可明显降低高糖诱导的细胞周期失调和KC凋亡[35]。相似地,本研究采用蛋白质印迹法和细胞免疫荧光法检测P62蛋白的表达,结果表明,高糖微环境下HaCaT中P62蛋白的表达显著增加,而NAC处理可明显抑制高糖微环境下HaCaT中P62蛋白的表达。这初步提示,活性氧可能是高糖微环境下KC中P62蛋白表达增加的上游机制。
本研究显示,在HaCaT中,高糖微环境促进P62蛋白的表达;敲减P62蛋白可明显促进KC迁移、增加KC运动性;高糖微环境下活性氧增加可能是P62表达增加的潜在机制。这在一定程度上揭示了P62在糖尿病创面愈合中的作用,为其作为难愈合创面治疗靶点提供了理论依据。
张喻平:实验操作、论文撰写;张琼、邓芳:数据整理、统计学分析;陈兵、张均辉、胡炯宇:研究指导、论文修改、经费支持所有作者均声明不存在利益冲突 -
参考文献
(35) [1] SyedFZ.Type 1 diabetes mellitus[J].Ann Intern Med,2022,175(3):ITC33-ITC48.DOI: 10.7326/AITC202203150. [2] TheocharidisG,ThomasBE,SarkarD,et al.Single cell transcriptomic landscape of diabetic foot ulcers[J].Nat Commun,2022,13(1):181.DOI: 10.1038/s41467-021-27801-8. [3] ShiMQ, DuZ, QiYC, et al. Wound microenvironment-responsive glucose consumption and hydrogen peroxide generation synergistic with azithromycin for diabetic wounds healing[J]. Theranostics, 2022, 12(6): 2658-2673. DOI: 10.7150/thno.64244. [4] KlionskyDJ, PetroniG, AmaravadiRK, et al. Autophagy in major human diseases[J]. EMBO J, 2021, 40(19): e108863. DOI: 10.15252/embj.2021108863. [5] RenHY, ZhaoF, ZhangQQ, et al. Autophagy and skin wound healing[J/OL]. Burns Trauma, 2022, 10:tkac003[2022-06-30]. https://pubmed.ncbi.nlm.nih.gov/35187180/.DOI: 10.1093/burnst/tkac003. [6] LevineB, KroemerG. Biological functions of autophagy genes: a disease perspective[J]. Cell, 2019, 176(1/2): 11-42. DOI: 10.1016/j.cell.2018.09.048. [7] FanXL, HuangTT, TongYD, et al. p62 works as a hub modulation in the ageing process[J]. Ageing Res Rev, 2022, 73: 101538.DOI: 10.1016/j.arr.2021.101538. [8] ChaoXJ,NiHM,DingWX. An unexpected tumor suppressor role of SQSTM1/p62 in liver tumorigenesis[J].Autophagy,2022,18(2):459-461.DOI: 10.1080/15548627.2021.2008693. [9] KarrasP,Riveiro-FalkenbachE,CañónE,et al.p62/SQSTM1 fuels melanoma progression by opposing mRNA decay of a selective set of pro-metastatic factors[J].Cancer Cell,2019,35(1):46-63.e10.DOI: 10.1016/j.ccell.2018.11.008. [10] Serrano-SaenzS, PalaciosC, Delgado-BellidoD, et al. PIM kinases mediate resistance of glioblastoma cells to TRAIL by a p62/SQSTM1-dependent mechanism[J]. Cell Death Dis, 2019, 10(2): 51. DOI: 10.1038/s41419-018-1293-3. [11] IslamMA,SooroMA,ZhangPH.Autophagic regulation of p62 is critical for cancer therapy[J].Int J Mol Sci,2018,19(5):1405.DOI: 10.3390/ijms19051405. [12] Sánchez-MartínP,KomatsuM.p62/SQSTM1-steering the cell through health and disease[J].J Cell Sci,2018,131(21): jcs.222836.DOI: 10.1242/jcs.222836. [13] MildenbergerJ, JohanssonI, SerginI, et al. N-3 PUFAs induce inflammatory tolerance by formation of KEAP1-containing SQSTM1/p62-bodies and activation of NFE2L2[J]. Autophagy, 2017, 13(10): 1664-1678.DOI: 10.1080/15548627.2017.1345411. [14] LiLF, ZhangJH, ZhangQ, et al. High glucose suppresses keratinocyte migration through the inhibition of p38 MAPK/autophagy pathway[J]. Front Physiol, 2019, 10: 24. DOI: 10.3389/fphys.2019.00024. [15] ZhangJH,LiLF,ZhangQ,et al.Microtubule-associated protein 4 phosphorylation regulates epidermal keratinocyte migration and proliferation[J].Int J Biol Sci,2019,15(9):1962-1976.DOI: 10.7150/ijbs.35440. [16] JiangT,LiQY,QiuJM,et al.Nanobiotechnology: applications in chronic wound healing[J].Int J Nanomedicine,2022,17:3125-3145.DOI: 10.2147/IJN.S372211. [17] ZhangJH, ZhangC, JiangXP, et al. Involvement of autophagy in hypoxia-BNIP3 signaling to promote epidermal keratinocyte migration[J]. Cell Death Dis, 2019, 10(3): 234. DOI: 10.1038/s41419-019-1473-9. [18] LiBM, TangHW, BianXW, et al. Calcium silicate accelerates cutaneous wound healing with enhanced re-epithelialization through EGF/EGFR/ERK-mediated promotion of epidermal stem cell functions[J/OL]. Burns Trauma, 2021, 9: tkab029[2022-06-30]. https://pubmed.ncbi.nlm.nih.gov/34604395/.DOI: 10.1093/burnst/tkab029. [19] ShawTJ,MartinP.Wound repair: a showcase for cell plasticity and migration[J].Curr Opin Cell Biol,2016,42:29-37.DOI: 10.1016/j.ceb.2016.04.001. [20] HuSC,LanCE.High-glucose environment disturbs the physiologic functions of keratinocytes: focusing on diabetic wound healing[J].J Dermatol Sci,2016,84(2):121-127.DOI: 10.1016/j.jdermsci.2016.07.008. [21] ParmarUM, JalgaonkarMP, KulkarniYA, et al. Autophagy-nutrient sensing pathways in diabetic complications[J]. Pharmacol Res, 2022, 184: 106408. DOI: 10.1016/j.phrs.202-2.106408. [22] MatooriS, VevesA, MooneyDJ. Advanced bandages for diabetic wound healing[J]. Sci Transl Med, 2021, 13(585): eabe4839.DOI: 10.1126/scitranslmed.abe4839. [23] NingSB, WangL. The multifunctional protein p62 and its mechanistic roles in cancers[J]. Curr Cancer Drug Targets, 2019, 19(6): 468-478. DOI: 10.2174/1568009618666181016164920. [24] TaoMM, LiuT, YouQD, et al. p62 as a therapeutic target for tumor[J]. Eur J Med Chem, 2020, 193: 112231. DOI: 10.1016/j.ejmech.2020.112231. [25] ZhangZ,CostaM.p62 functions as a signal hub in metal carcinogenesis[J].Semin Cancer Biol,2021,76:267-278.DOI: 10.1016/j.semcancer.2021.04.014. [26] YouZY,JiangWX,QinLY,et al.Requirement for p62 acetylation in the aggregation of ubiquitylated proteins under nutrient stress[J].Nat Commun,2019,10(1):5792.DOI: 10.1038/s41467-019-13718-w. [27] GradaA, Otero-VinasM, Prieto-CastrilloF, et al. Research techniques made simple: analysis of collective cell migration using the wound healing assay[J]. J Invest Dermatol, 2017, 137(2): e11-e16.DOI: 10.1016/j.jid.2016.11.020. [28] 张均辉,张琼,贾杰只,等.低氧条件下B淋巴细胞瘤-2/腺病毒E1B19 000相互作用蛋白3对人真皮微血管内皮细胞迁移和运动性的影响及其机制[J].中华烧伤杂志,2021,37(1):9-16.DOI: 10.3760/cma.j.cn501120-20200927-00425. [29] DengLL, DuCZ, SongPY, et al. The role of oxidative stress and antioxidants in diabetic wound healing[J]. Oxid Med Cell Longev, 2021, 2021: 8852759. DOI: 10.1155/2021/8852759. [30] WeiXR, LiMX, ZhengZJ, et al. Senescence in chronic wounds and potential targeted therapies[J/OL]. Burns Trauma, 2022, 10: tkab045[2022-06-30]. https://pubmed.ncbi.nlm.nih.gov/35187179/.DOI: 10.1093/burnst/tkab045. [31] QiuJX, ZhangT, ZhuXY, et al. Hyperoside induces breast cancer cells apoptosis via ROS-mediated NF-κB signaling pathway[J]. Int J Mol Sci, 2020, 21(1): 131. DOI: 10.3390/ijms21010131. [32] HeinkeL. Mitochondrial ROS drive cell cycle progression[J]. Nat Rev Mol Cell Biol, 2022, 23(9): 581. DOI: 10.1038/s41580-022-00523-5. [33] ShenY,YangJJ,ZhaoJ,et al.The switch from ER stress-induced apoptosis to autophagy via ROS-mediated JNK/p62 signals: a survival mechanism in methotrexate-resistant choriocarcinoma cells[J].Exp Cell Res,2015,334(2):207-218.DOI: 10.1016/j.yexcr.2015.04.010. [34] BalszuweitF, MenacherG, SchmidtA, et al. Protective effects of the thiol compounds GSH and NAC against sulfur mustard toxicity in a human keratinocyte cell line[J]. Toxicol Lett, 2016, 244: 35-43. DOI: 10.1016/j.toxlet.2015.09.002. [35] RizwanH, PalS, SabnamS, et al. High glucose augments ROS generation regulates mitochondrial dysfunction and apoptosis via stress signalling cascades in keratinocytes[J]. Life Sci, 2020, 241: 117148. DOI: 10.1016/j.lfs.2019.117148. -
1 蛋白质印迹法检测正常对照组和高糖培养3个时间点组HaCaT中P62蛋白表达
注:GAPDH为3-磷酸甘油醛脱氢酶;条带上方1、2、3、4分别指示正常对照组、高糖24 h组、高糖48 h组、高糖72 h组
2 正常对照组和高糖组HaCaT培养48 h的P62蛋白表达 异硫氰酸荧光素-4',6-二脒基-2-苯基吲哚×630。2A、2B、2C.分别为正常对照组P62染色、细胞核染色、P62与细胞核染色重叠图片,细胞核完整,P62蛋白表达较少,主要分布于细胞质;2D、2E、2F.分别为高糖组P62染色、细胞核染色、P62与细胞核染色重叠图片,细胞核完整,P62蛋白表达较图2A多
注:P62阳性染色为绿色,细胞核阳性染色为蓝色
3 蛋白质印迹法检测转染siRNA后4组HaCaT中P62蛋白表达
注:GAPDH为3-磷酸甘油醛脱氢酶,siRNA为小干扰RNA;条带上方1、2、3、4分别指示阴性对照siRNA组、P62-siRNA-1组、P62-siRNA-2组、P62-siRNA-3组
4 蛋白质印迹法检测高糖微环境下siRNA对4组HaCaT转染后72 h的P62蛋白表达的作用
注:GAPDH为3-磷酸甘油醛脱氢酶,siRNA为小干扰RNA;条带上方1、2、3、4分别指示正常糖+阴性对照siRNA组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组
5 4组HaCaT处理完成后划痕后各时间点细胞划痕面积情况 倒置光学显微镜×100。5A、5B、5C、5D.分别为正常糖+阴性对照小干扰RNA(siRNA)组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组划痕后0 h(即刻),划痕情况相近;5E、5F、5G、5H.分别为正常糖+阴性对照siRNA组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组划痕后24 h,图5F划痕面积明显小于图5E,图5G划痕面积明显大于图5E,图5H划痕面积明显小于图5G
6 活细胞工作站观察4组HaCaT 3 h内的运动范围。6A、6B、6C、6D.分别为正常糖+阴性对照siRNA组、正常糖+P62-siRNA组、高糖+阴性对照siRNA组、高糖+P62-siRNA组,其中图6B、6D运动范围分别较图6A、6C明显增大
注:siRNA为小干扰RNA;细胞运动起点均为坐标(0,0),运动终点为位于4个象限中的圆点,连接两者之间的曲线为细胞运动轨迹
7 蛋白质印迹法检测高糖微环境下NAC对3组HaCaT培养48 h的P62蛋白表达的影响
注:GAPDH为3-磷酸甘油醛脱氢酶,PBS为磷酸缓冲盐溶液,NAC为N-乙酰半胱氨酸;条带上方1、2、3分别指示正常糖+PBS组、高糖+PBS组、高糖+NAC组
8 3组HaCaT培养48 h的P62蛋白表达 异硫氰酸荧光素-4',6-二脒基-2-苯基吲哚×630。8A、8B、8C.分别为正常糖+PBS组P62染色、细胞核染色、P62与细胞核染色重叠图片,细胞核完整,P62蛋白表达较少,主要分布于细胞质;8D、8E、8F.分别为高糖+PBS组P62染色、细胞核染色、P62与细胞核染色重叠图片,细胞核完整,P62蛋白表达较图8A多;8G、8H、8I.分别为高糖+NAC组P62染色、细胞核染色、P62与细胞核染色重叠图片,细胞核完整,P62蛋白表达较图8D少
注:P62阳性染色为绿色,细胞核阳性染色为蓝色;PBS为磷酸缓冲盐溶液,NAC为N-乙酰半胱氨酸
-


 下载:
下载:
 下载:
下载:

计量
- 文章访问数: 1235
- HTML全文浏览量: 116
- PDF下载量: 39
- 被引次数: 0
