



Research advances on the effects of RNA N6-methyladenosine modification in the relevant pathophysiological processes of wound repair
-
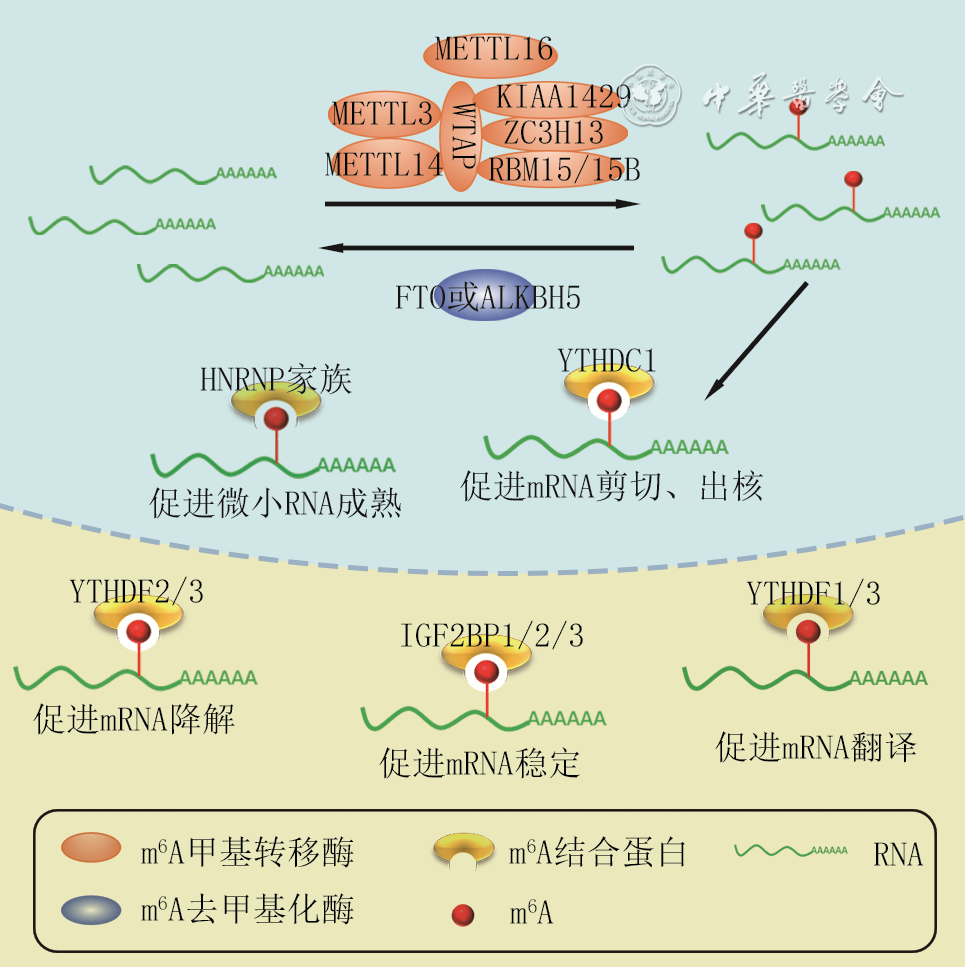
摘要: N6-甲基腺嘌呤(m6A)作为一种转录后修饰广泛存在于真核生物中,该种修饰受到甲基转移酶和去甲基化酶的动态可逆性调控,并通过m6A结合蛋白参与调节生物学效应。近来研究表明,m6A可参与胚胎皮肤形态发生、创面修复以及炎症反应、血管生成和纤维化等病理生理过程。该文概述了m6A及其相关蛋白在创面修复相关病理生理进程中的作用,以期为创面修复的治疗策略提供新的理论依据。Abstract: N6-methyladenosine (m6A) exists widely in eukaryotes as a post-transcriptional modification. This modification is dynamically and reversibly regulated by methyltransferases and demethylases, and is involved in regulating biological effects through m6A binding proteins. Recent studies have elucidated that m6A is involved in embryonic skin morphogenesis, wound repair, and pathophysiological processes such as inflammatory response, angiogenesis, and fibrosis. This review summarizes the role of m6A and its related proteins in the related pathophysiological processes of wound repair, so as to provide a new theoretical basis for the treatment strategy of wound repair.
-
参考文献
(40) [1] HuangH,WengH,ChenJ.m6A modification in coding and non-coding RNAs: roles and therapeutic implications in cancer[J].Cancer Cell,2020,37(3):270-288.DOI: 10.1016/j.ccell.2020.02.004. [2] HePC,HeC.m6A RNA methylation: from mechanisms to therapeutic potential[J].EMBO J,2021,40(3):e105977.DOI: 10.15252/embj.2020105977. [3] DominissiniD,Moshitch-MoshkovitzS,SchwartzS,et al.Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq[J].Nature,2012,485(7397):201-206.DOI: 10.1038/nature11112. [4] MeyerKD,SaletoreY,ZumboP,et al.Comprehensive analysis of mRNA methylation reveals enrichment in 3' UTRs and near stop codons[J].Cell,2012,149(7):1635-1646.DOI: 10.1016/j.cell.2012.05.003. [5] FangX,LiM,YuT,et al.Reversible N6-methyladenosine of RNA: the regulatory mechanisms on gene expression and implications in physiology and pathology[J].Genes Dis,2020,7(4):585-597.DOI: 10.1016/j.gendis.2020.06.011. [6] ShiH,WeiJ,HeC.Where, when, and how: context-dependent functions of RNA methylation writers, readers, and erasers[J].Mol Cell,2019,74(4):640-650.DOI: 10.1016/j.molcel.2019.04.025. [7] YangY,HsuPJ,ChenYS,et al.Dynamic transcriptomic m6A decoration: writers, erasers, readers and functions in RNA metabolism[J].Cell Res,2018,28(6):616-624.DOI: 10.1038/s41422-018-0040-8. [8] XiL,CarrollT,MatosI,et al.m6A RNA methylation impacts fate choices during skin morphogenesis[J].Elife,2020,9:e56980.DOI: 10.7554/eLife.56980. [9] VeltriA,LangC,LienWH.Concise review: Wnt signaling pathways in skin development and epidermal stem cells[J].Stem Cells,2018,36(1):22-35.DOI: 10.1002/stem.2723. [10] LeeJ,WuY,HaradaBT,et al.N6-methyladenosine modification of lncRNA Pvt1 governs epidermal stemness[J].EMBO J,2021,40(8):e106276.DOI: 10.15252/embj.2020106276. [11] RodriguesM,KosaricN,BonhamCA,et al.Wound healing: a cellular perspective[J].Physiol Rev,2019,99(1):665-706.DOI: 10.1152/physrev.00067.2017. [12] WangJ,YanS,LuH,et al.METTL3 attenuates LPS-induced inflammatory response in macrophages via NF-κB signaling pathway[J].Mediators Inflamm,2019,2019:3120391.DOI: 10.1155/2019/3120391. [13] ZhangY,GuX,LiD,et al.METTL3 regulates osteoblast differentiation and inflammatory response via Smad signaling and MAPK signaling[J].Int J Mol Sci,2019,21(1):199.DOI: 10.3390/ijms21010199. [14] ZongX,ZhaoJ,WangH,et al.Mettl3 deficiency sustains long-chain fatty acid absorption through suppressing Traf6-dependent inflammation response[J].J Immunol,2019,202(2):567-578.DOI: 10.4049/jimmunol.1801151. [15] FengZ,LiQ,MengR,et al.METTL3 regulates alternative splicing of MyD88 upon the lipopolysaccharide-induced inflammatory response in human dental pulp cells[J].J Cell Mol Med,2018,22(5):2558-2568.DOI: 10.1111/jcmm.13491. [16] JianD,WangY,JianL,et al.METTL14 aggravates endothelial inflammation and atherosclerosis by increasing FOXO1 N6-methyladeosine modifications[J].Theranostics,2020,10(20):8939-8956.DOI: 10.7150/thno.45178. [17] DuJ,LiaoW,LiuW,et al.N6-adenosine methylation of Socs1 mRNA is required to sustain the negative feedback control of macrophage activation[J].Dev Cell,2020,55(6):737-753.e7.DOI: 10.1016/j.devcel.2020.10.023. [18] HouJ,ZhangH,LiuJ,et al.YTHDF2 reduction fuels inflammation and vascular abnormalization in hepatocellular carcinoma[J].Mol Cancer,2019,18(1):163.DOI: 10.1186/s12943-019-1082-3. [19] MapperleyC,van de LagemaatLN,LawsonH,et al.The mRNA m6A reader YTHDF2 suppresses proinflammatory pathways and sustains hematopoietic stem cell function[J].J Exp Med,2021,218(3):e20200829.DOI: 10.1084/jem.20200829. [20] YuR,LiQ,FengZ,et al.m6A reader YTHDF2 regulates LPS-induced inflammatory response[J].Int J Mol Sci,2019,20(6):1323.DOI: 10.3390/ijms20061323. [21] ZhengL,TangX,LuM,et al.microRNA-421-3p prevents inflammatory response in cerebral ischemia/reperfusion injury through targeting m6A reader YTHDF1 to inhibit p65 mRNA translation[J].Int Immunopharmacol,2020,88:106937.DOI: 10.1016/j.intimp.2020.106937. [22] ZhaoJ,HanDX,WangCB,et al.Zbtb7b suppresses aseptic inflammation by regulating m6A modification of IL6 mRNA[J].Biochem Biophys Res Commun,2020,530(1):336-341.DOI: 10.1016/j.bbrc.2020.07.011. [23] WuC,ChenW,HeJ,et al.Interplay of m6A and H3K27 trimethylation restrains inflammation during bacterial infection[J].Sci Adv,2020,6(34):eaba0647.DOI: 10.1126/sciadv.aba0647. [24] ChenL,ChengL,ChenT,et al.Macrophage polarization in skin wound healing: progress in biology and therapeutics[J].J Shanghai Jiaotong Univ (Sci),2021,27:264-280.DOI: 10.1007/s12204-021-2276-6. [25] GuX,ZhangY,LiD,et al.N6-methyladenosine demethylase FTO promotes M1 and M2 macrophage activation[J].Cell Signal,2020,69:109553.DOI: 10.1016/j.cellsig.2020.109553. [26] LiuY,LiuZ,TangH,et al.The N6-methyladenosine (m6A)-forming enzyme METTL3 facilitates M1 macrophage polarization through the methylation of STAT1 mRNA[J].Am J Physiol Cell Physiol,2019,317(4):C762-C775.DOI: 10.1152/ajpcell.00212.2019. [27] WangQ,ChenC,DingQ,et al.METTL3-mediated m6A modification of HDGF mRNA promotes gastric cancer progression and has prognostic significance[J].Gut,2020,69(7):1193-1205.DOI: 10.1136/gutjnl-2019-319639. [28] YangZ,WangT,WuD,et al.RNA N6-methyladenosine reader IGF2BP3 regulates cell cycle and angiogenesis in colon cancer[J].J Exp Clin Cancer Res,2020,39(1):203.DOI: 10.1186/s13046-020-01714-8. [29] WangLJ,XueY,LiH,et al.Wilms' tumour 1-associating protein inhibits endothelial cell angiogenesis by m6A-dependent epigenetic silencing of desmoplakin in brain arteriovenous malformation[J].J Cell Mol Med,2020,24(9):4981-4991.DOI: 10.1111/jcmm.15101. [30] MathiyalaganP,AdamiakM,MayourianJ,et al.FTO-dependent N6-methyladenosine regulates cardiac function during remodeling and repair[J].Circulation,2019,139(4):518-532.DOI: 10.1161/CIRCULATIONAHA.118.033794. [31] ShanK,ZhouRM,XiangJ,et al.FTO regulates ocular angiogenesis via m6A-YTHDF2-dependent mechanism[J].Exp Eye Res,2020,197:108107.DOI: 10.1016/j.exer.2020.108107. [32] WangLJ,XueY,HuoR,et al.N6-methyladenosine methyltransferase METTL3 affects the phenotype of cerebral arteriovenous malformation via modulating Notch signaling pathway[J].J Biomed Sci,2020,27(1):62.DOI: 10.1186/s12929-020-00655-w. [33] YaoMD,JiangQ,MaY,et al.Role of METTL3-dependent N6-methyladenosine mRNA modification in the promotion of angiogenesis[J].Mol Ther,2020,28(10):2191-2202.DOI: 10.1016/j.ymthe.2020.07.022. [34] LiuSY,WuJJ,ChenZH,et al.The m6A RNA modification modulates gene expression and fibrosis-related pathways in hypertrophic scar[J].Front Cell Dev Biol,2021,9:748703.DOI: 10.3389/fcell.2021.748703. [35] LiT,ZhuangY,YangW,et al.Silencing of METTL3 attenuates cardiac fibrosis induced by myocardial infarction via inhibiting the activation of cardiac fibroblasts[J].FASEB J,2021,35(2):e21162.DOI: 10.1096/fj.201903169R. [36] LinX,ChaiG,WuY,et al.RNA m6A methylation regulates the epithelial mesenchymal transition of cancer cells and translation of Snail[J].Nat Commun,2019,10(1):2065.DOI: 10.1038/s41467-019-09865-9. [37] TangB,YangY,KangM,et al.m6A demethylase ALKBH5 inhibits pancreatic cancer tumorigenesis by decreasing WIF-1 RNA methylation and mediating Wnt signaling[J].Mol Cancer,2020,19(1):3.DOI: 10.1186/s12943-019-1128-6. [38] NingY,ChenJ,ShiY,et al.Genistein ameliorates renal fibrosis through regulation snail via m6A RNA demethylase ALKBH5[J].Front Pharmacol,2020,11:579265.DOI: 10.3389/fphar.2020.579265. [39] HanB,ChuC,SuX,et al.N6-methyladenosine-dependent primary microRNA-126 processing activated PI3K-AKT-mTOR pathway drove the development of pulmonary fibrosis induced by nanoscale carbon black particles in rats[J].Nanotoxicology,2020,14(1):1-20.DOI: 10.1080/17435390.2019.1661041. [40] LiuP,ZhangB,ChenZ,et al.m6A-induced lncRNA MALAT1 aggravates renal fibrogenesis in obstructive nephropathy through the miR-145/FAK pathway[J].Aging (Albany NY),2020,12(6):5280-5299.DOI: 10.18632/aging.102950. -

 下载:
下载:

图(1)
计量
- 文章访问数: 252
- HTML全文浏览量: 98
- PDF下载量: 13
- 被引次数: 0
