



Effects of tumor necrosis factor-alpha/extracellular signal-regulated kinase pathway on migration ability of HaCaT cells and full-thickness skin defects in mice
-
摘要:
目的 探讨肿瘤坏死因子α(TNF-α)/胞外信号调节激酶(ERK)途径对HaCaT细胞迁移能力及小鼠全层皮肤缺损的影响。 方法 采用实验研究方法。按照随机数字表法(下同)将HaCaT细胞分为常氧组和低氧(氧气体积分数为1%,下同)条件下培养的低氧组。培养24 h后,采用微阵列置信度分析软件SAM 4.01筛选出2组细胞的显著差异表达基因,通过京都基因和基因组百科全书对信号通路中每条基因数目的显著性进行分析以筛选差异显著的信号通路,样本数为3。另取HaCaT细胞,在低氧条件下培养0(即刻)、3、6、12、24 h,采用酶联免疫吸附测定(ELISA)法检测细胞分泌TNF-α的水平,样本数为5。另取HaCaT细胞,分为常氧组、单纯低氧组和加入FR180204(一种ERK抑制剂)并置于低氧条件下培养的低氧+抑制剂组,培养3、6、12、24 h,采用划痕试验检测细胞的迁移能力,样本数为12。另取HaCaT细胞,在低氧条件下培养0、3、6、12、24 h,采用蛋白质印迹法检测细胞中磷酸化核因子κB(p-NF-κB)、磷酸化p38(p-p38)、磷酸化ERK1/2(p-ERK1/2)、神经钙黏素和上皮钙黏素的表达,样本数为3。取64只6~8周龄雄性BALB/c小鼠,在小鼠背部制作全层皮肤缺损创面模型后,将其分为空白对照组和用FR180204处理的抑制剂组,每组32只,分别作相应处理。伤后0、3、6、9、12、15 d,观察创面情况并计算其愈合率(样本数为8)。伤后1、3、6、15 d,采用苏木精-伊红染色观察创面中新生血管生成、炎症细胞浸润和表皮新生情况,采用Masson染色观察创面中胶原沉积情况,采用蛋白质印迹法检测创面组织中p-NF-κB、p-p38、p-ERK1/2、神经钙黏素和上皮钙黏素的表达(样本数为6),采用免疫组织化学法检测创面中Ki67阳性细胞数和血管内皮生长因子(VEGF)吸光度值(样本数为5),采用ELISA法检测创面组织中白细胞介素6(IL-6)、IL-10、IL-1β和CCL20的蛋白表达(样本数为6)。对数据行单因素方差分析、重复测量方差分析、析因设计方差分析、Tukey检验、LSD检验和独立样本t检验。 结果 培养24 h后,与常氧组相比,低氧组细胞中上调了7 667个基因,下调了7 174个基因;在前述差异表达基因中,TNF-α信号通路有显著的变化(P<0.05)且基因数目多。低氧条件下,与0 h的(1.9±0.3)pg/mL相比,TNF-α表达量仅在细胞培养24 h[(11.1±2.1)pg/mL]时显著增加(P<0.05)。与常氧组相比,单纯低氧组细胞培养6、12、24 h的迁移能力均明显增强(t值分别为2.27、4.65、4.67,P<0.05);与单纯低氧组相比,低氧+抑制剂组细胞培养3、6、12、24 h的迁移能力均明显减弱(t值分别为2.43、3.06、4.62、8.14,P<0.05)。低氧条件下,与培养0 h相比,p-NF-κB、p-ERK1/2、神经钙黏素表达量在细胞培养12、24 h时均明显升高(P<0.05),p-p38表达量在细胞培养3、6、12、24 h时均明显升高(P<0.05),上皮钙黏素表达量在细胞培养6、12、24 h时均明显降低(P<0.05);其中,p-ERK1/2、p-NF-κB和上皮钙黏素在细胞中的表达量呈时间依赖性。与空白对照组相比,抑制剂组小鼠伤后3、6、9、12、15 d创面愈合率均明显降低(P<0.05);抑制剂组小鼠创缘周围在伤后3、6、15 d有更多炎症细胞浸润,尤其在伤后15 d,创面中可见大量组织坏死且新生表皮层不连续,同时胶原合成和新生血管减少;抑制剂组小鼠创面中p-NF-κB表达量在伤后3、6 d均明显降低(t值分别为3.26、4.26,P<0.05)但在伤后15 d明显升高(t=3.25,P<0.05),p-p38和神经钙黏素表达量在伤后1、3、6 d均明显降低(t值分别为4.89、2.98、3.98,9.51、11.69、4.10,P<0.05),p-ERK1/2表达量在伤后1、3、6、15 d均明显降低(t值分别为26.69、3.63、5.12、5.14,P<0.05),上皮钙黏素表达量在伤后1 d明显降低(t=20.67,P<0.05)但在伤后6 d明显增加(t=2.90,P<0.05);抑制剂组小鼠创面中Ki67阳性细胞数和VEGF的吸光度值在伤后3、6、15 d均明显减少/降低(t值分别为4.20、7.35、3.34,4.14、3.20、3.73,P<0.05);抑制剂组小鼠创面组织中IL-10表达量在伤后6 d明显降低(t=2.92,P<0.05),IL-6表达量在伤后6 d明显增加(t=2.73,P<0.05),IL-1β表达量在伤后15 d显著增加(t=3.46,P<0.05),CCL20表达量在伤后1、6 d均明显降低(t值分别为3.96、2.63,P<0.05)但在伤后15 d显著增加(t=3.68,P<0.05)。 结论 TNF-α/ERK途径可促进HaCaT细胞迁移并通过影响小鼠全层皮肤缺损创面中炎症因子和趋化因子的表达调控创面愈合。 -
关键词:
- 伤口愈合 /
- 肿瘤坏死因子α /
- 皮肤 /
- 胞外信号调节激酶通路 /
- 表皮细胞迁移
Abstract:Objective To investigate the effects of tumor necrosis factor-alpha (TNF-α)/extracellular signal-regulated kinase (ERK) pathway on the migration ability of HaCaT cells and full-thickness skin defects in mice. Methods The experimental research method was adopted. According to the random number table (the same below), HaCaT cells were divided into the normal oxygen group and the hypoxia group cultured under hypoxia (with oxygen volume fraction of 1%, the same below) condition. After 24 hours of culture, the significantly differentially expressed genes between the 2 groups were screened using the microarray confidence analysis software SAM4.01. The significance of the number of each gene in the signaling pathway was analyzed through the Kyoto encyclopedia of genes and genomes to screen the significantly differentially signaling pathways (n=3). HaCaT cells were cultured for 0 (immediately), 3, 6, 12, and 24 h under hypoxia condition. The secretion level of TNF-α was detected by enzyme-linked immunosorbent assay (ELISA), and the number of samples was 5. HaCaT cells were divided into normal oxygen group, hypoxia alone group, and hypoxia+inhibitor group cultured with FR180204 (an ERK inhibitor) and under hypoxia condition. The cells were cultured for 3, 6, 12, and 24 h. The migration ability of the cells was detected by scratch test (n=12). The expressions of phosphorylated nuclear factor kappa B (p-NF-κB), phosphorylated p38 (p-p38), phosphorylated ERK1/2 (p-ERK1/2), N-cadherin, and E-cadherin in HaCaT cells were detected by Western blotting under hypoxic condition for 0, 3, 6, 12, and 24 h (n=3). Sixty-four BALB/c male mice aged 6 to 8 weeks were used to make a full-thickness skin defect wound model on the dorsum of the mice. The mice were divided into the blank control group and the inhibitor group treated with FR180204, with 32 mice in each group being treated accordingly. On post injury day (PID) 0, 3, 6, 9, 12, and 15, the wound conditions of mice were observed and the healing rate was calculated (n=8). On PID 1, 3, 6, and 15, hematoxylin-eosin staining was used to observe neovascularization, inflammatory cell infiltration, and epidermal regeneration on wound, Masson staining was used to observe collagen deposition on wound, the expressions of p-NF-κB, p-p38, p-ERK12, N-cadherin, and E-cadherin in wound tissue were detected by Western blotting (n=6), the number of Ki67 positive cells and the absorbance value of vascular endothelial growth factor (VEGF) were detected by immunohistochemistry (n=5), the protein expressions of interleukin 6 (IL-6), IL-10, IL-1β, and CCL20 in wound tissue were detected by ELISA (n=6). Data were statistically analyzed with one-way analysis of variance, analysis of variance for repeated measurement, factorial design analysis of variance, Tukey test, least significant difference test, and independent sample t test. Results After 24 hours of culture, compared with normal oxygen group, 7 667 genes were up-regulated and 7 174 genes were down-regulated in cells in hypoxic group. Among the above differentially expressed genes, the TNF-α signaling pathway had significant change (P<0.05) with large number of genes. Under hypoxia condition, the expression of TNF-α at 24 h of cell culture was (11.1±2.1) pg/mL, which was significantly higher than (1.9±0.3) pg/mL at 0 h (P<0.05). Compared with normal oxygen group, the migration ability of cells in hypoxia alone group was significantly enhanced at 6, 12, and 24 h of cell culture (with t values of 2.27, 4.65, and 4.67, respectively, P<0.05). Compared with hypoxia alone group, the migration ability of cells in hypoxia+inhibitor group was significantly decreased at 3, 6, 12, and 24 h of cell culture (with t values of 2.43, 3.06, 4.62, and 8.14, respectively, P<0.05). Under hypoxia condition, the expressions of p-NF-κB, p-ERK1/2, and N-cadherin were increased significantly at 12 and 24 h of cell culture compared with 0 h of culture (P<0.05), the expression of p-p38 was significantly increased at 3, 6, 12, and 24 h of cell culture (P<0.05), the expression of E-cadherin was significantly decreased at 6, 12, and 24 h of cell culture (P<0.05), the expression of p-ERK1/2, p-NF-κB, and E-cadherin was time-dependent. Compared with blank control group, on PID 3, 6, 9, 12, and 15, the wound healing rate of mice in inhibitor group was significantly decreased (P<0.05); there were more inflammatory cell infiltration around the wound edge of mice in inhibitor group on PID 3, 6, and 15, especially on PID 15, a large number of tissue necrosis and discontinuous new epidermal layer were observed on the wound surface, and collagen synthesis and new blood vessels were reduced; the expression of p-NF-κB in the wound of mice in inhibitor group was significantly decreased on PID 3 and 6 (with t values of 3.26 and 4.26, respectively, P<0.05) but significantly increased on PID 15 (t=3.25, P<0.05), the expressions of p-p38 and N-cadherin were significantly decreased on PID 1, 3, and 6 (with t values of 4.89, 2.98, 3.98, 9.51, 11.69, and 4.10, respectively, P<0.05), the expression of p-ERK1/2 was significantly decreased on PID 1, 3, 6, and 15 (with t values of 26.69, 3.63, 5.12, and 5.14, respectively, P<0.05), the expression of E-cadherin was significantly decreased on PID 1 (t=20.67, P<0.05) but significantly increased on PID 6 (t=2.90, P<0.05); the number of Ki67 positive cells and absorbance value of VEGF of wound in inhibitor group were significantly decreased on PID 3, 6, and 15 (with t values of 4.20, 7.35, 3.34, 4.14, 3.20, and 3.73, respectively, P<0.05); the expression of IL-10 in the wound tissue of the inhibitor group was significantly decreased on PID 6 (t=2.92, P<0.05), the expression of IL-6 was significantly increased on PID 6 (t=2.73, P<0.05), the expression of IL-1β was significantly increased on PID 15 (t=3.46, P<0.05), and CCL20 expression levels were significantly decreased on PID 1 and 6 (with t values of 3.96 and 2.63, respectively, P<0.05) but significantly increased on PID 15 (t=3.68, P<0.05). Conclusions The TNF-α/ERK pathway can promote the migration of HaCaT cells, and regulate the healing of full-thickness skin defect wounds in mice by affecting the expression of inflammatory cytokines and chemokines. -
参考文献
(33) [1] WilkinsonHN,HardmanMJ.Wound healing: cellular mechanisms and pathological outcomes[J].Open Biol,2020,10(9):200223.DOI: 10.1098/rsob.200223. [2] ZhangJ,YanY,LiY,et al.Topical effect of benzalkonium bromide on wound healing and potential cellular and molecular mechanisms[J].Int Wound J,2021,18(5):566-576.DOI: 10.1111/iwj.13555. [3] LiaoF,ChenL,LuoP,et al.PC4 serves as a negative regulator of skin wound healing in mice[J/OL].Burns Trauma,2020,8:tkaa010[2022-10-19].https://pubmed.ncbi.nlm.nih.gov/32373645/.DOI: 10.1093/burnst/tkaa010. [4] LiL,HeY,ZhaoM,et al.Collective cell migration: Implications for wound healing and cancer invasion[J/OL].Burns Trauma,2013,1(1):21-26[2022-10-19].https://pubmed.ncbi.nlm.nih.gov/27574618/.DOI: 10.4103/2321-3868.113331. [5] KiwanukaE,HacklF,CatersonEJ,et al.CCN2 is transiently expressed by keratinocytes during re-epithelialization and regulates keratinocyte migration in vitro by the ras-MEK-ERK signaling pathway[J].J Surg Res,2013,185(2):e109-119.DOI: 10.1016/j.jss.2013.05.065. [6] RuthenborgRJ,BanJJ,WazirA,et al.Regulation of wound healing and fibrosis by hypoxia and hypoxia-inducible factor-1[J].Mol Cells,2014,37(9):637-643.DOI: 10.14348/molcells.2014.0150. [7] SternD,CuiH.Crafting Polymeric and peptidic hydrogels for improved wound healing[J].Adv Healthc Mater,2019,8(9):e1900104.DOI: 10.1002/adhm.201900104. [8] ZhangYM,ZhangZQ,LiuYY,et al.Requirement of Gαi1/3-Gab1 signaling complex for keratinocyte growth factor-induced PI3K-AKT-mTORC1 activation[J].J Invest Dermatol,2015,135(1):181-191.DOI: 10.1038/jid.2014.326. [9] ChenX,ZhouX,MaoTC,et al.Effect of microtubule-associated protein-4 on epidermal cell migration under different oxygen concentrations[J].J Dermatol,2016,43(6):674-681.DOI: 10.1111/1346-8138.13192. [10] ChenX,ZhouX,ShiX,et al.MAP4 regulates Tctex-1 and promotes the migration of epidermal cells in hypoxia[J].Exp Dermatol,2018,27(11):1210-1215.DOI: 10.1111/exd.13763. [11] SchremlS,SzeimiesRM,PrantlL,et al.Oxygen in acute and chronic wound healing[J].Br J Dermatol,2010,163(2):257-268.DOI: 10.1111/j.1365-2133.2010.09804.x. [12] TaiG,TaiM,ZhaoM.Electrically stimulated cell migration and its contribution to wound healing[J/OL].Burns Trauma,2018,6:20[2022-10-19].https://pubmed.ncbi.nlm.nih.gov/30003115/.DOI: 10.1186/s41038-018-0123-2. [13] FuX.Wound healing center establishment and new technology application in improving the wound healing quality in China[J/OL].Burns Trauma,2020,8:tkaa038[2022-10-19].https://pubmed.ncbi.nlm.nih.gov/33134399/.DOI: 10.1093/burnst/tkaa038. [14] 黄跃生.调控生物电场与氧微环境促进创面再生修复[J].中华烧伤杂志,2021,37(1):5-8.DOI: 10.3760/cma.j.cn501120-20201123-00492. [15] RamaswamyP,GoswamiK,Dalavaikodihalli NanjaiahN,et al.TNF-α mediated MEK-ERK signaling in invasion with putative network involving NF-κB and STAT-6: a new perspective in glioma[J].Cell Biol Int,2019,43(11):1257-1266.DOI: 10.1002/cbin.11125. [16] BalkwillF.Tumour necrosis factor and cancer[J].Nat Rev Cancer,2009,9(5):361-371.DOI: 10.1038/nrc2628. [17] LuoJL,MaedaS,HsuLC,et al.Inhibition of NF-kappaB in cancer cells converts inflammation- induced tumor growth mediated by TNFalpha to TRAIL-mediated tumor regression[J].Cancer Cell,2004,6(3):297-305.DOI: 10.1016/j.ccr.2004.08.012. [18] AshcroftGS,JeongMJ,AshworthJJ,et al.Tumor necrosis factor-alpha (TNF-α) is a therapeutic target for impaired cutaneous wound healing[J].Wound Repair Regen,2012,20(1):38-49.DOI: 10.1111/j.1524-475X.2011.00748.x. [19] YenYH,PuCM,LiuCW,et al.Curcumin accelerates cutaneous wound healing via multiple biological actions: the involvement of TNF-α, MMP-9, α-SMA, and collagen[J].Int Wound J,2018,15(4):605-617.DOI: 10.1111/iwj.12904. [20] YanC,GrimmWA,GarnerWL,et al.Epithelial to mesenchymal transition in human skin wound healing is induced by tumor necrosis factor-alpha through bone morphogenic protein-2[J].Am J Pathol,2010,176(5):2247-2258.DOI: 10.2353/ajpath.2010.090048. [21] HaY,LeeWH,JeongJ,et al.Pyropia yezoensis extract suppresses IFN-gamma- and TNF-alpha-induced proinflammatory chemokine production in HaCaT cells via the down-regulation of NF-κB[J].Nutrients,2020,12(5)DOI: 10.3390/nu12051238. [22] WenLJ,HuXL,LiCY,et al.Myosin 1b promotes migration, invasion and glycolysis in cervical cancer via ERK/HIF-1α pathway[J].Am J Transl Res,2021,13(11):12536-12548. [23] 张灿,张琼,张均辉,等.组蛋白脱乙酰酶6抑制剂Tubastatin A对人皮肤成纤维细胞增殖和运动的影响及其分子机制[J].中华烧伤杂志,2021,37(9):853-859.DOI: 10.3760/cma.j.cn501120-20200519-00274. [24] LiB,TangH,BianX,et al.Calcium silicate accelerates cutaneous wound healing with enhanced re-epithelialization through EGF/EGFR/ERK-mediated promotion of epidermal stem cell functions[J/OL].Burns Trauma,2021,9:tkab029[2022-10-19].https://pubmed.ncbi.nlm.nih.gov/34604395/.DOI: 10.1093/burnst/tkab029. [25] BrunerHC,DerksenP.Loss of E-cadherin-dependent cell-cell adhesion and the development and progression of Cancer[J].Cold Spring Harb Perspect Biol,2018,10(3):a029330.DOI: 10.1101/cshperspect.a029330. [26] NiemanMT,PrudoffRS,JohnsonKR,et al.N-cadherin promotes motility in human breast cancer cells regardless of their E-cadherin expression[J].J Cell Biol,1999,147(3):631-644.DOI: 10.1083/jcb.147.3.631. [27] WalkerA,FreiR,LawsonKR.The cytoplasmic domain of N-cadherin modulates MMP-9 induction in oral squamous carcinoma cells[J].Int J Oncol,2014,45(4):1699-1706.DOI: 10.3892/ijo.2014.2549. [28] ZhangH,SunP,WangYL,et al.MiR-214 promotes proliferation and inhibits apoptosis of oral cancer cells through MAPK/ERK signaling pathway[J].Eur Rev Med Pharmacol Sci,2020,24(7):3710-3716.DOI: 10.26355/eurrev_202004_20834. [29] KhanNM,HaseebA,AnsariMY,et al.Wogonin, a plant derived small molecule, exerts potent anti-inflammatory and chondroprotective effects through the activation of ROS/ERK/Nrf2 signaling pathways in human Osteoarthritis chondrocytes[J].Free Radic Biol Med,2017,106:288-301.DOI: 10.1016/j.freeradbiomed.2017.02.041. [30] DongQ,JieY,MaJ,et al.Renal tubular cell death and inflammation response are regulated by the MAPK-ERK-CREB signaling pathway under hypoxia-reoxygenation injury[J].J Recept Signal Transduct Res,2019,39(5-6):383-391.DOI: 10.1080/10799893.2019.1698050. [31] FurueK,ItoT,TsujiG,et al.The CCL20 and CCR6 axis in psoriasis[J].Scand J Immunol,2020,91(3):e12846.DOI: 10.1111/sji.12846. [32] DuttaP,TaA,ThakurBK,et al.Biphasic Ccl20 regulation by Toll-like receptor 9 through the activation of ERK-AP-1 and non-canonical NF-κB signaling pathways[J].Biochim Biophys Acta Gen Subj,2017,1861(1 Pt A):3365-3377.DOI: 10.1016/j.bbagen.2016.08.019. [33] 刘中阳,程旭,张景霞,等.Vγ4T细胞在应用雷帕霉素的小鼠全层皮肤缺损创面愈合障碍中的作用及其机制[J].中华烧伤与创面修复杂志,2022,38(5):462-470.DOI: 10.3760/cma.j.cn501120-20201209-00523. -

1 低氧条件(较常氧条件)下培养24 h后HaCaT细胞差异表达基因分析(样本数为3)
注:低氧条件指氧气体积分数为1%;TNF为肿瘤坏死因子,PI3K为磷脂酰肌醇-3-激酶,Akt为蛋白激酶B,EGFR为表皮生长因子受体;P值为低氧组与常氧组相比所得;椭圆大小表示基因数目多少

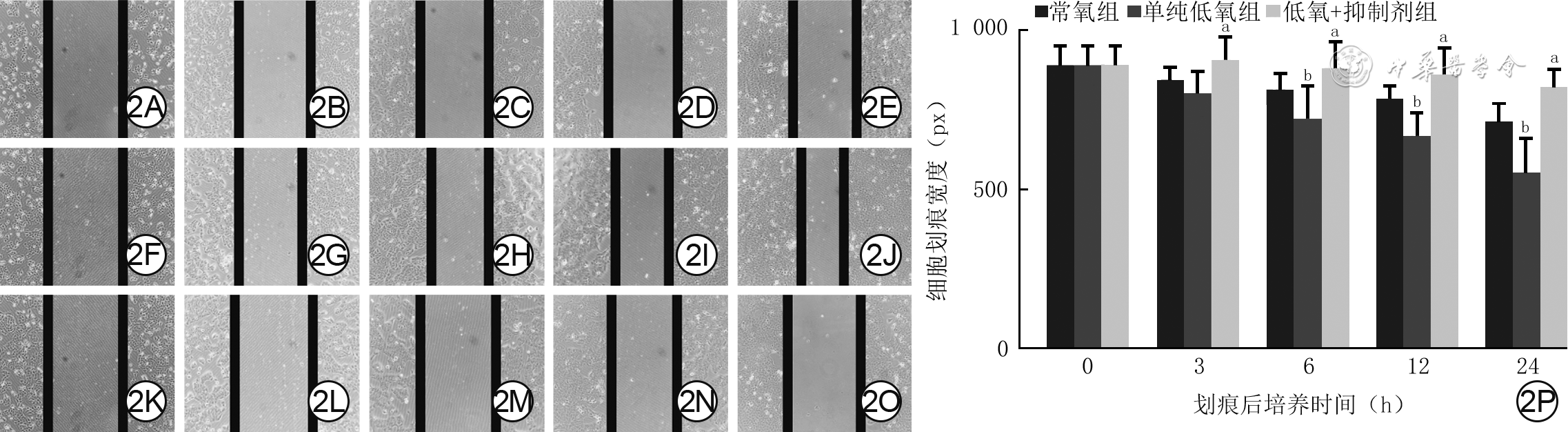
2 3组HaCaT细胞不同培养时间点的迁移情况。2A、2B、2C、2D、2E,2F、2G、2H、2I、2J,2K、2L、2M、2N、2O.分别为常氧组、单纯低氧组、低氧+抑制剂组细胞在划痕后培养0(即刻)、3、6、12、24 h情况,图2I、2J的划痕宽度分别较图2D、2E明显变窄,图2L、2M、2N、2O的划痕宽度分别较图2G、2H、2I、2J明显变宽;2P.不同时间点细胞划痕宽度的条图(样本数为12,
注:低氧条件指氧气体积分数为1%;抑制剂组所使用抑制剂为FR180204;划痕宽度反映细胞迁移能力

3 低氧条件下培养各时间点HaCaT细胞相关蛋白表达情况。3A.各蛋白的条带图;3B.各蛋白的条图(样本数为3,
注:低氧条件指氧气体积分数为1%;p-ERK1/2为磷酸化胞外信号调节激酶1/2,GAPDH为3-磷酸甘油醛脱氢酶;条带图上方的1、2、3、4、5分别指细胞培养0(即刻)、3、6、12、24 h;将培养0 h的各蛋白表达量设为1,其余各指标蛋白表达量与之相比为相对表达量

4 2组全层皮肤缺损小鼠伤后不同时间点创面愈合情况。4A、4B、4C、4D、4E、4F.分别为空白对照组伤后0、3、6、9、12、15 d的创面情况;4G、4H、4I、4J、4K、4L.分别为抑制剂组伤后0、3、6、9、12、15 d的创面情况,其中图4H、4I、4J、4K、4L的创面面积均分别明显大于图4B、4C、4D、4E、4F;4M.2组小鼠伤后不同时间点的创面愈合率比较(样本数为8,
注:抑制剂组所使用抑制剂为FR180204

5 2组全层皮肤缺损小鼠伤后不同时间点创面组织中相关蛋白表达。5A.条带图,条带图上方1、3、5、7为空白对照组伤后1、3、6、15 d创面情况,条带图上方2、4、6、8为抑制剂组伤后1、3、6、15 d创面情况;5B.条图(样本数为6,
注:抑制剂组所使用抑制剂为FR180204;p-ERK1/2为磷酸化胞外信号调节激酶1/2;GAPDH指3-磷酸甘油醛脱氢酶;将空白对照组各蛋白的表达量设为1,抑制剂组各蛋白的表达量与之相比为相对表达量
表1 2组全层皮肤缺损小鼠伤后不同时间点创面组织中细胞因子表达比较(pg/mL,
组别 样本数 白细胞介素10 白细胞介素1β 1 d 3 d 6 d 15 d 1 d 3 d 6 d 15 d 空白对照组 6 90±6 94±7 87±7 73±9 40.0±5.2 42.2±2.1 41.2±2.9 31.2±1.4 抑制剂组 6 89±8 90±6 68±8 64±7 41.3±3.4 43.5±4.2 41.5±4.6 37.8±4.1 t值 0.34 0.90 2.92 0.81 0.55 0.70 0.15 3.46 P值 0.744 0.394 0.019 0.443 0.593 0.503 0.883 0.013 注:抑制剂组所使用抑制剂为FR180204  下载: 导出CSV
下载: 导出CSV
-

 下载:
下载:






计量
- 文章访问数: 2893
- HTML全文浏览量: 128
- PDF下载量: 40
- 被引次数: 0
