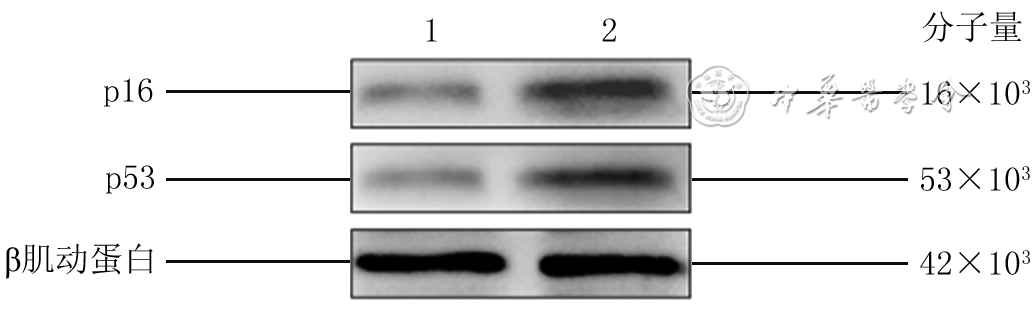
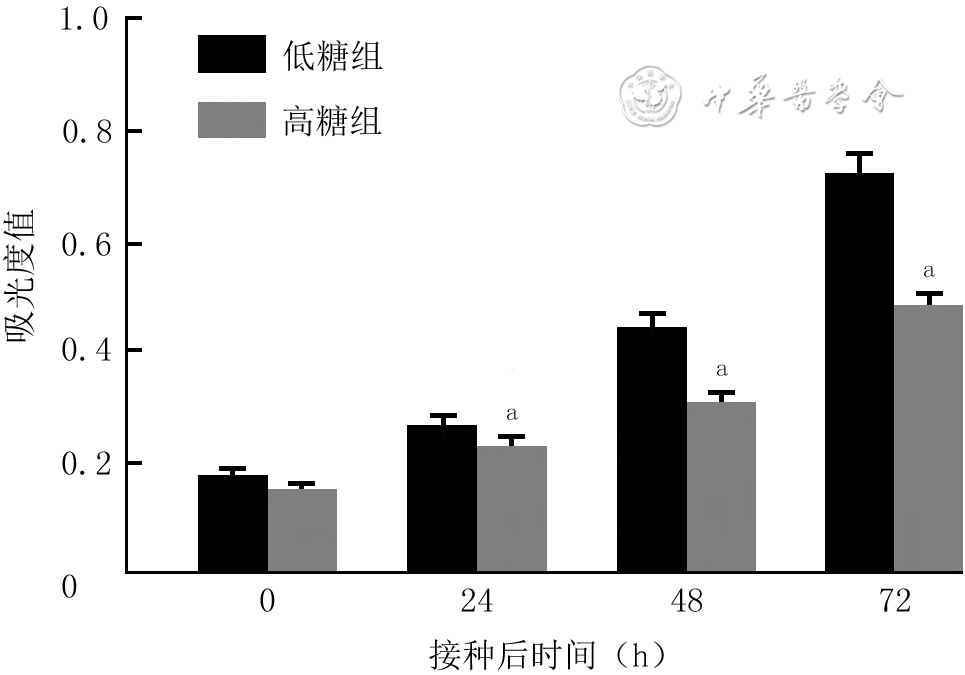
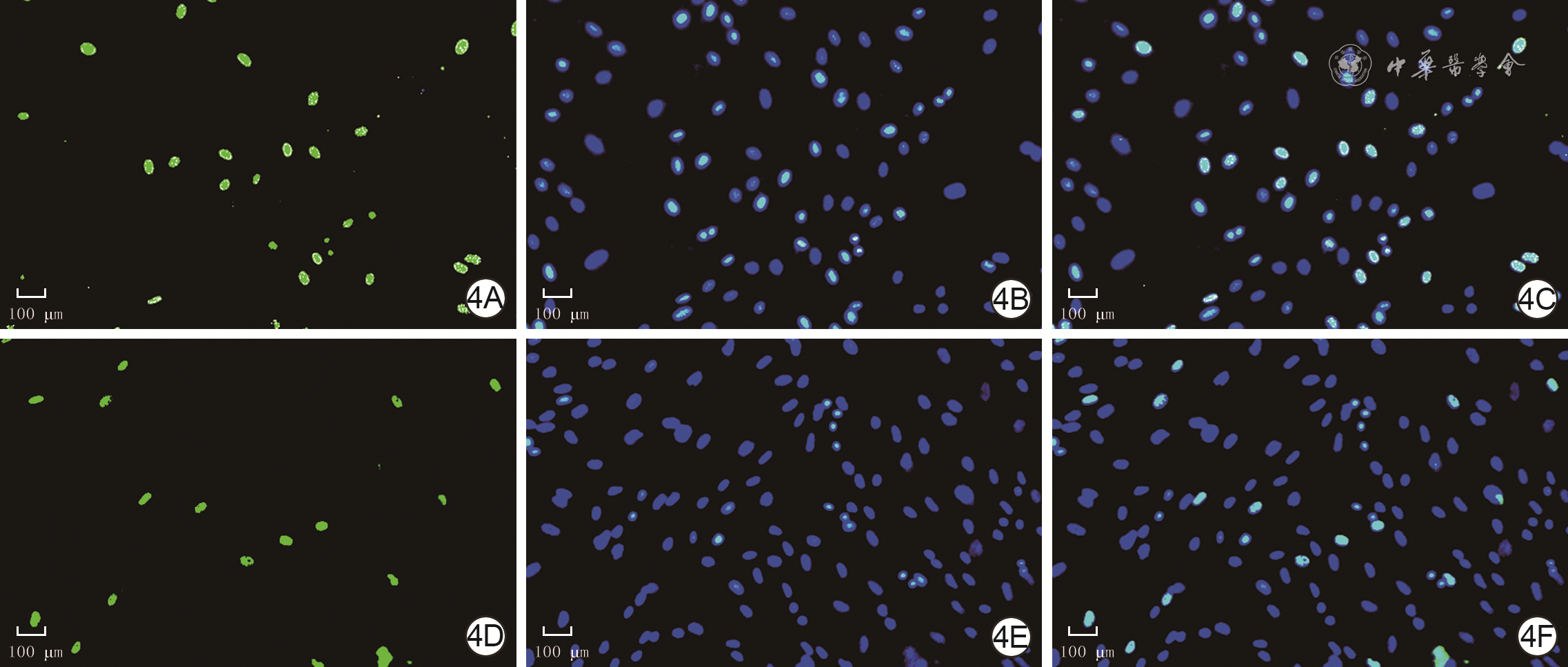
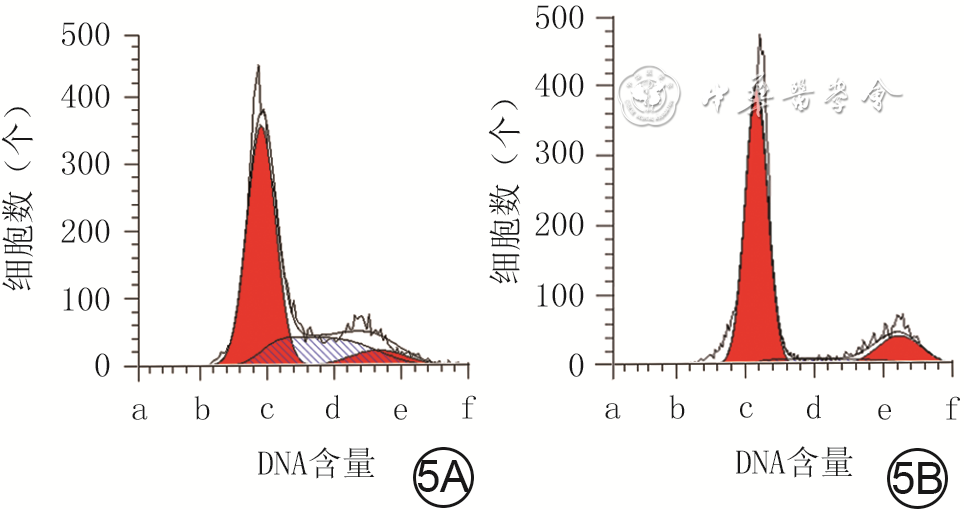
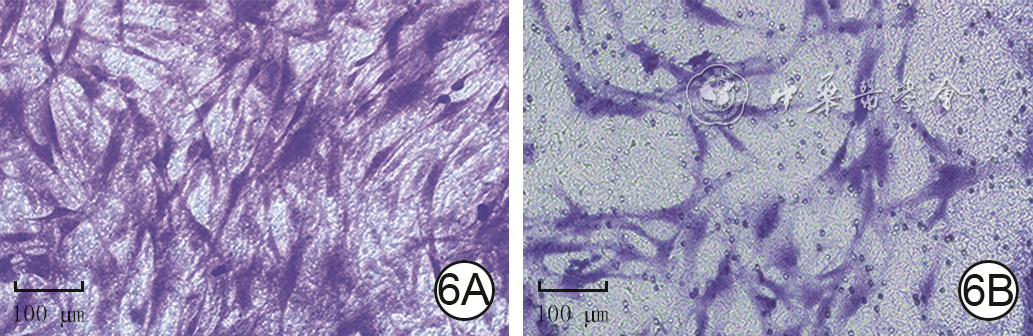
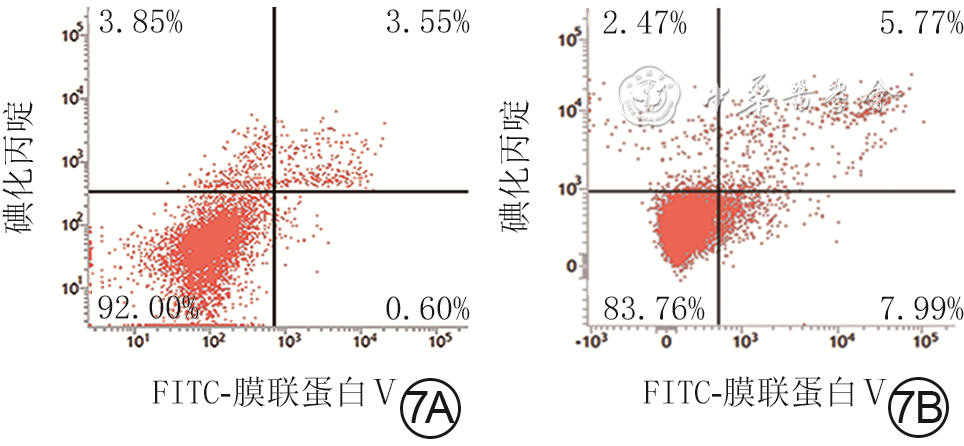
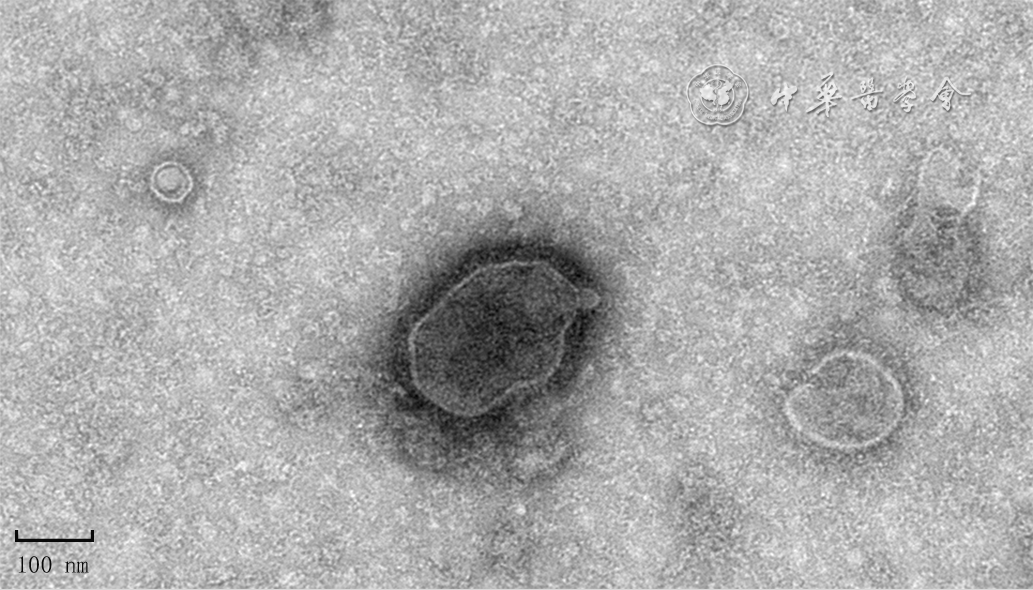
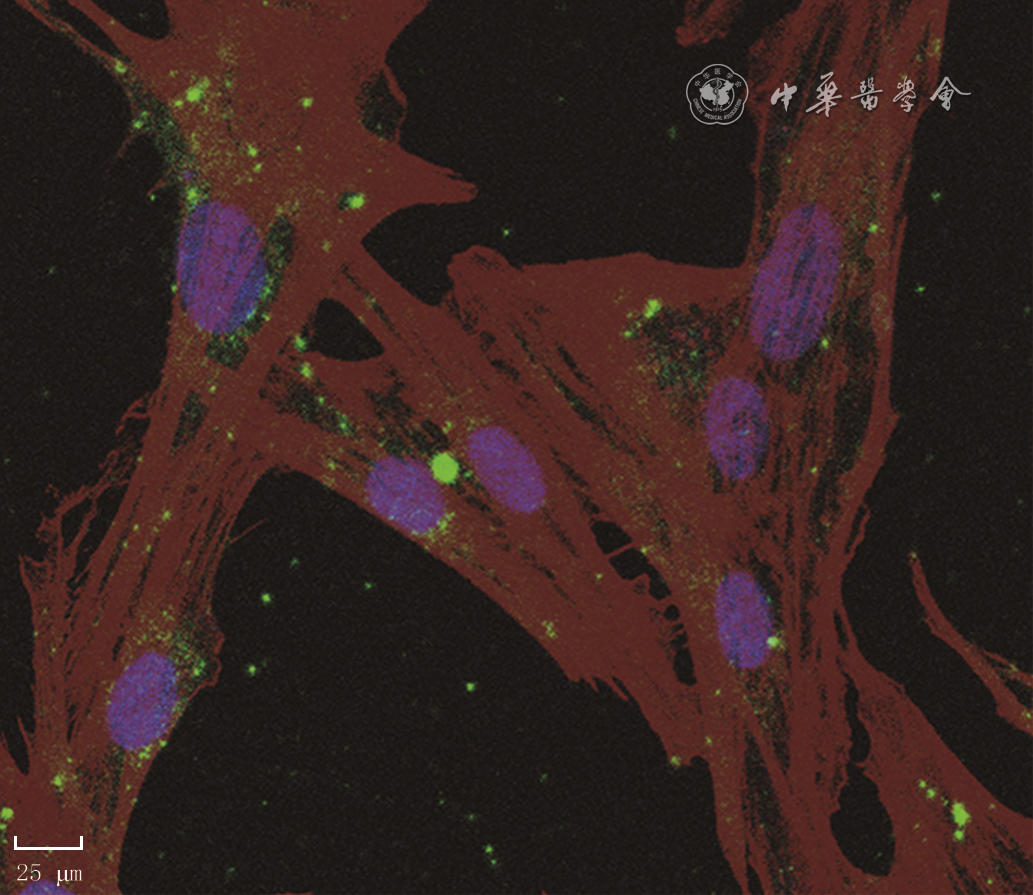
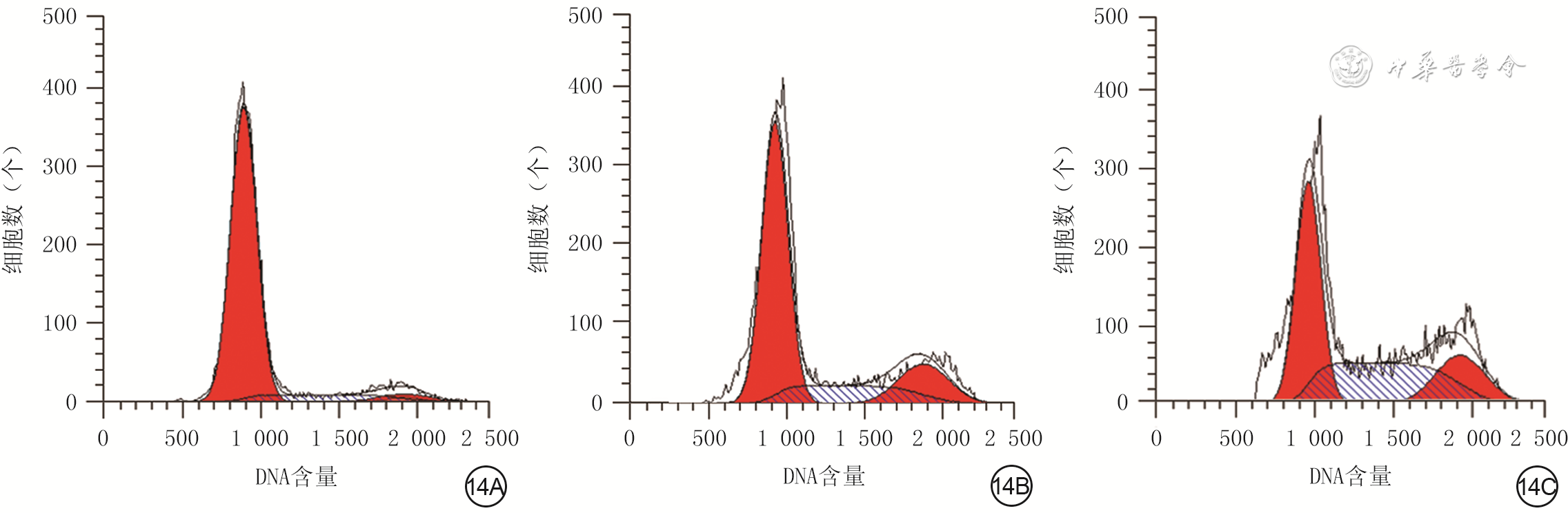
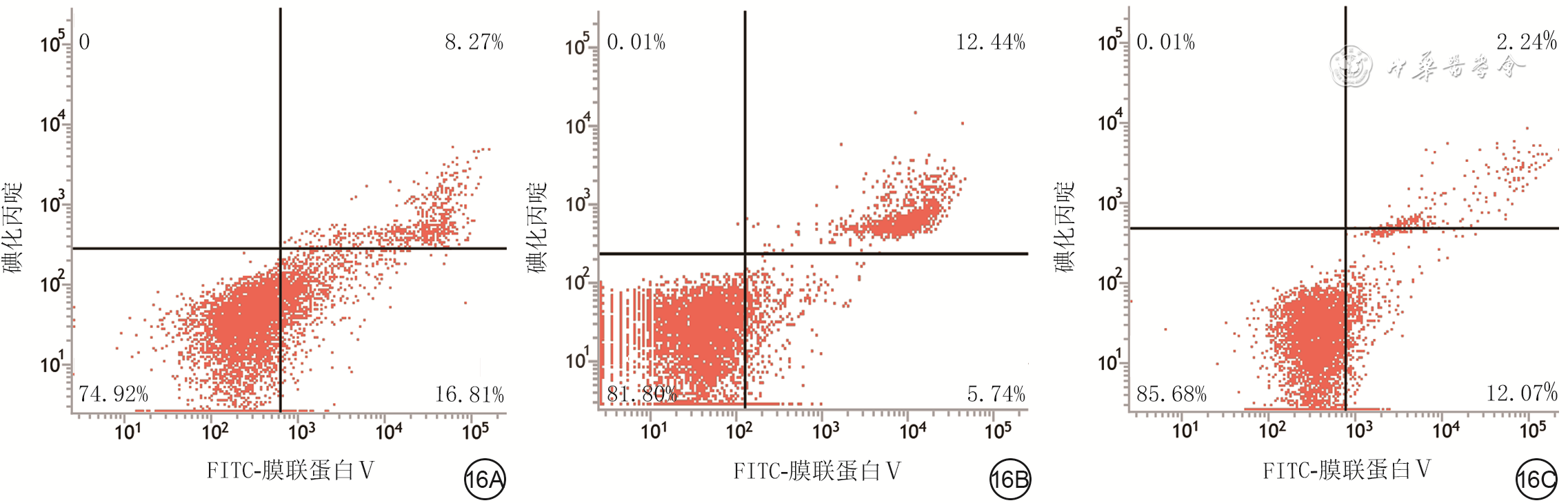
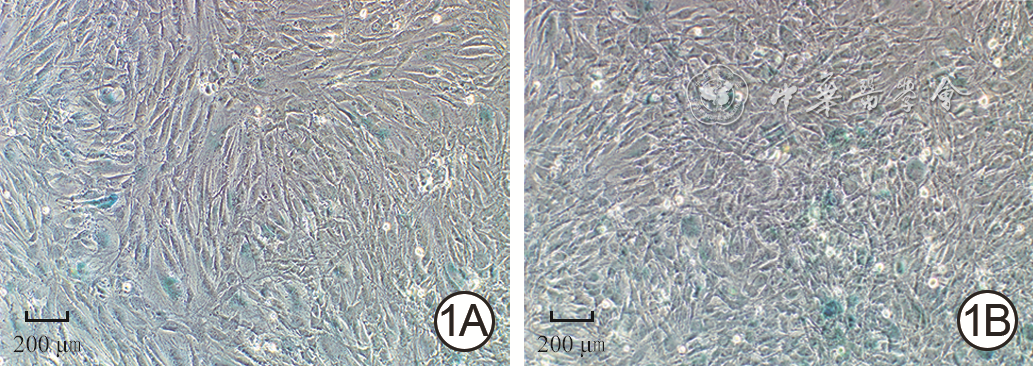
| Citation: | Su JL,Ma K,Zhang CP,et al.Effect of human decidua mesenchymal stem cells-derived exosomes on the function of high glucose-induced senescent human dermal fibroblasts and its possible mechanism[J].Chin J Burns Wounds,2022,38(2):170-183.DOI: 10.3760/cma.j.cn501120-20210925-00330.

|
| [1] |
LiB, BianX, HuW, et al. Regenerative and protective effects of calcium silicate on senescent fibroblasts induced by high glucose[J]. Wound Repair Regen,2020,28(3):315-325. DOI: 10.1111/wrr.12794.
|
| [2] |
LiM,ZhaoY,HaoH,et al.Umbilical cord-derived mesenchymal stromal cell-conditioned medium exerts in vitro antiaging effects in human fibroblasts[J].Cytotherapy,2017,19(3):371-383.DOI: 10.1016/j.jcyt.2016.12.001.
|
| [3] |
RaniS,RyanAE,GriffinMD,et al.Mesenchymal stem cell-derived extracellular vesicles: toward cell-free therapeutic applications[J].Mol Ther,2015,23(5):812-823.DOI: 10.1038/mt.2015.44.
|
| [4] |
NikfarjamS,RezaieJ,ZolbaninNM,et al.Mesenchymal stem cell derived-exo somes: a modern approach in translational medicine[J].J Transl Med,2020,18(1):449.DOI: 10.1186/s12967-020-02622-3.
|
| [5] |
KomakiM,NumataY,MoriokaC,et al.Exosomes of human placenta-derived mesenchymal stem cells stimulate angiogenesis[J].Stem Cell Res Ther,2017,8(1):219.DOI: 10.1186/s13287-017-0660-9.
|
| [6] |
ZhaoB,ZhangY,HanS,et al.Exosomes derived from human amniotic epithelial cells accelerate wound healing and inhibit scar formation[J].J Mol Histol,2017,48(2):121-132.DOI: 10.1007/s10735-017-9711-x.
|
| [7] |
DashBC,XuZ,LinL,et al.Stem cells and engineered scaffolds for regenerative wound healing[J].Bioengineering (Basel),2018, 5(1):23. DOI: 10.3390/bioengineering5010023.
|
| [8] |
LermanOZ,GalianoRD,ArmourM,et al.Cellular dysfunction in the diabetic fibroblast: impairment in migration, vascular endothelial growth factor production, and response to hypoxia[J].Am J Pathol,2003,162(1):303-312.DOI: 10.1016/S0002-9440(10)63821-7.
|
| [9] |
ThomasK,KiwitM,KernerW.Glucose concentration in human subcutaneous adipose tissue: comparison between forearm and abdomen[J].Exp Clin Endocrinol Diabetes,1998,106(6):465-469.DOI: 10.1055/s-0029-1212017.
|
| [10] |
WeiQ,WangY,MaK,et al.Extracellular vesicles from human umbilical cord mesenchymal stem cells facilitate diabetic wound healing through miR-17-5p-mediated enhancement of angiogenesis[J/OL].Stem Cell Rev Rep,2021(2021-05-04)[2021-10-13]. https://pubmed.ncbi.nlm.nih.gov/33942217/. DOI:10.1007/s12015-021-10176-0. [published online ahead of print].
|
| [11] |
ChengL,ZhangK,WuS,et al.Focus on mesenchymal stem cell-derived exosomes: opportunities and challenges in cell-free therapy[J].Stem Cells Int,2017,2017:6305295.DOI: 10.1155/2017/6305295.
|
| [12] |
LuK,LiHY,YangK,et al.Exosomes as potential alternatives to stem cell therapy for intervertebral disc degeneration: in-vitro study on exosomes in interaction of nucleus pulposus cells and bone marrow mesenchymal stem cells[J].Stem Cell Res Ther,2017,8(1):108.DOI: 10.1186/s13287-017-0563-9.
|
| [13] |
Casado-DíazA,Quesada-GómezJM,DoradoG.Extracellular vesicles derived from mesenchymal stem cells (MSC) in regenerative medicine: applications in skin wound healing[J].Front Bioeng Biotechnol,2020,8:146.DOI: 10.3389/fbioe.2020.00146.
|
| [14] |
HuY,RaoSS,WangZX,et al.Exosomes from human umbilical cord blood accelerate cutaneous wound healing through miR-21-3p-mediated promotion of angiogenesis and fibroblast function[J].Theranostics,2018,8(1):169-184.DOI: 10.7150/thno.21234.
|
| [15] |
PomattoM,GaiC,NegroF,et al.Differential therapeutic effect of extracellular vesicles derived by bone marrow and adipose mesenchymal stem cells on wound healing of diabetic ulcers and correlation to their cargoes[J].Int J Mol Sci,2021, 22(8):3851. DOI: 10.3390/ijms22083851.
|
| [16] |
WangC,WangM,XuT,et al.Engineering bioactive self-healing antibacterial exosomes hydrogel for promoting chronic diabetic wound healing and complete skin regeneration[J].Theranostics,2019,9(1):65-76.DOI: 10.7150/thno.29766.
|
| [17] |
McBrideJD,Rodriguez-MenocalL,GuzmanW,et al.Bone marrow mesenchymal stem cell-derived CD63+ exosomes transport Wnt3a exteriorly and enhance dermal fibroblast proliferation, migration, and angiogenesis in vitro[J].Stem Cells Dev,2017,26(19):1384-1398.DOI: 10.1089/scd.2017.0087.
|
| [18] |
YangJ,ChenZ,PanD,et al.Umbilical cord-derived mesenchymal stem cell-derived exosomes combined pluronic F127 hydrogel promote chronic diabetic wound healing and complete skin regeneration[J].Int J Nanomedicine,2020,15:5911-5926.DOI: 10.2147/IJN.S249129.
|
| [19] |
ZhangW,BaiX,ZhaoB,et al.Cell-free therapy based on adipose tissue stem cell-derived exosomes promotes wound healing via the PI3K/Akt signaling pathway[J].Exp Cell Res,2018,370(2):333-342.DOI: 10.1016/j.yexcr.2018.06.035.
|
| [20] |
FangS,XuC,ZhangY,et al.Umbilical cord-derived mesenchymal stem cell-derived exosomal microRNAs suppress myofibroblast differentiation by inhibiting the transforming growth factor-β/SMAD2 pathway during wound healing[J].Stem Cells Transl Med,2016,5(10):1425-1439.DOI: 10.5966/sctm.2015-0367.
|
| [21] |
HuL,WangJ,ZhouX,et al.Exosomes derived from human adipose mensenchymal stem cells accelerates cutaneous wound healing via optimizing the characteristics of fibroblasts[J].Sci Rep,2016,6:32993.DOI: 10.1038/srep32993.
|
| [22] |
WiklanderOP, NordinJZ, O'LoughlinA,et al.Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting[J].J Extracell Vesicles,2015,4:26316.DOI: 10.3402/jev.v4.26316.
|
| [23] |
YangD,ZhangW,ZhangH,et al.Progress, opportunity, and perspective on exosome isolation - efforts for efficient exosome-based theranostics[J].Theranostics,2020,10(8):3684-3707.DOI: 10.7150/thno.41580.
|
| [24] |
DingM,WangC,LuX,et al.Comparison of commercial exosome isolation kits for circulating exosomal microRNA profiling[J].Anal Bioanal Chem,2018,410(16):3805-3814.DOI: 10.1007/s00216-018-1052-4.
|
| [25] |
KusumaGD,CarthewJ,LimR,et al.Effect of the microenvironment on mesenchymal stem cell paracrine signaling: opportunities to engineer the therapeutic effect[J].Stem Cells Dev,2017,26(9):617-631.DOI: 10.1089/scd.2016.0349.
|
| [26] |
WuD,KangL,TianJ,et al.Exosomes derived from bone mesenchymal stem cells with the stimulation of Fe3O4 nanoparticles and static magnetic field enhance wound healing through upregulated miR-21-5p[J].Int J Nanomedicine,2020,15:7979-7993.DOI: 10.2147/IJN.S275650.
|
| [27] |
YangH,LinJ,JiangJ,et al.miR-20b-5p functions as tumor suppressor microRNA by targeting cyclinD1 in colon cancer[J].Cell Cycle,2020,19(21):2939-2954.DOI: 10.1080/15384101.2020.1829824.
|
| [28] |
DimitrovaN,GochevaV,BhutkarA,et al.Stromal expression of miR-143/145 promotes neoangiogenesis in lung cancer development[J].Cancer Discov,2016,6(2):188-201.DOI: 10.1158/2159-8290.CD-15-0854.
|
| [29] |
DimitrovaN,GochevaV,BhutkarA,et al.Stromal expression of miR-143/145 promotes neoangiogenesis in lung cancer development[J].Cancer Discov,2016,6(2):188-201.DOI: 10.1158/2159-8290.CD-15-0854.
|
| [30] |
LawsonJ,DickmanC,MacLellanS,et al.Selective secretion of microRNAs from lung cancer cells via extracellular vesicles promotes CAMK1D-mediated tube formation in endothelial cells[J].Oncotarget,2017,8(48):83913-83924.DOI: 10.18632/oncotarget.19996.
|
| [31] |
GrasC,RatusznyD,HadamitzkyC,et al.miR-145 contributes to hypertrophic scarring of the skin by inducing myofibroblast activity[J].Mol Med,2015,21(1):296-304.DOI: 10.2119/molmed.2014.00172.
|
| [32] |
ChaiC,WuH,WangB,et al.MicroRNA-498 promotes proliferation and migration by targeting the tumor suppressor PTEN in breast cancer cells[J].Carcinogenesis,2018,39(9):1185-1196.DOI: 10.1093/carcin/bgy092.
|
| [33] |
DuanXM,LiuXN,LiYX,et al.MicroRNA-498 promotes proliferation, migration, and invasion of prostate cancer cells and decreases radiation sensitivity by targeting PTEN[J].Kaohsiung J Med Sci,2019,35(11):659-671.DOI: 10.1002/kjm2.12108.
|
| [34] |
LiJ,ZhangF,LiH,et al.Circ_0010220-mediated miR-503-5p/CDCA4 axis contributes to osteosarcoma progression tumorigenesis[J].Gene,2020,763:145068.DOI: 10.1016/j.gene.2020.145068.
|
| [35] |
MarkopoulosGS,RoupakiaE,TokamaniM,et al.Senescence-associated microRNAs target cell cycle regulatory genes in normal human lung fibroblasts[J].Exp Gerontol,2017,96:110-122.DOI: 10.1016/j.exger.2017.06.017.
|
| [36] |
JiangL,ZhaoZ,ZhengL,et al.Downregulation of miR-503 promotes ESCC cell proliferation, migration, and invasion by targeting cyclin D1[J].Genomics Proteomics Bioinformatics,2017,15(3):208-217.DOI: 10.1016/j.gpb.2017.04.003.
|
| [37] |
BovyN,BlommeB,FrèresP,et al.Endothelial exosomes contribute to the antitumor response during breast cancer neoadjuvant chemotherapy via microRNA transfer[J].Oncotarget,2015,6(12):10253-10266.DOI: 10.18632/oncotarget.3520.
|
| [38] |
HouSQ,OuyangM,BrandmaierA,et al.PTEN in the maintenance of genome integrity: From DNA replication to chromosome segregation[J].Bioessays,2017,39(10):10. DOI: 10.1002/bies.201700082.
|
| [39] |
LiB,LuanS,ChenJ,et al.The MSC-derived exosomal incRNA H19 promotes wound healing in diabetic foot ulcers by upregulating PTEN via microRNA-152-3p[J].Mol Ther Nucleic Acids,2020,19:814-826.DOI: 10.1016/j.omtn.2019.11.034.
|
| [40] |
LongJ,OuC,XiaH,et al.MiR-503 inhibited cell proliferation of human breast cancer cells by suppressing CCND1 expression[J].Tumour Biol,2015,36(11):8697-8702.DOI: 10.1007/s13277-015-3623-8.
|

Figures(16) / Tables(2)
Copyright © Chinese Journal of Burns京ICP备07035254号-14
E-mail:shaoshangzazhi@163.com
Supported by:
Beijing Renhe Information Technology Co. Ltd






 DownLoad:
DownLoad: