



Effects and mechanism of age on the stiffness and the fibrotic phenotype of fibroblasts of human hypertrophic scar
-
摘要:
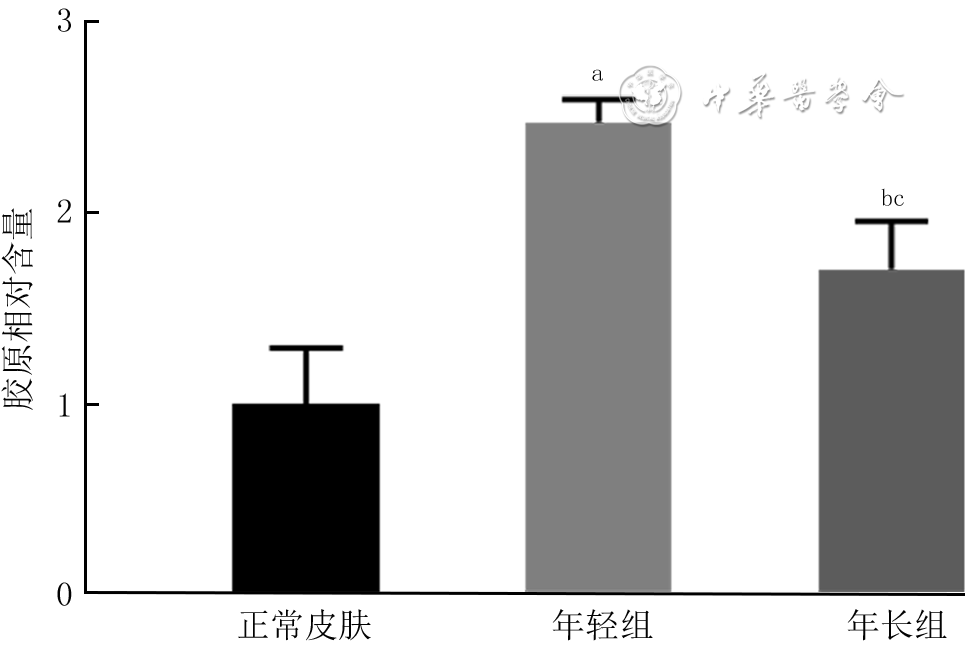
目的 探讨年龄对人增生性瘢痕硬度和成纤维细胞(Fb)纤维化表型的影响及其可能的分子机制。 方法 采用实验研究方法。收集2020年1—6月解放军总医院第四医学中心烧伤整形外科收治的10例瘢痕患者(男4例、女6例)手术切除的增生性瘢痕组织和10例患者(男5例、女5例,年龄7~41岁)手术后剩余的正常全层皮肤组织。根据患者年龄,将6例患者[(10.7±1.6)岁]瘢痕组织纳入年轻组,将4例患者[(40.0±2.2)岁]瘢痕组织纳入年长组。对正常皮肤和2组瘢痕组织,行苏木精-伊红(HE)染色观察组织形态,行Masson染色观察胶原形态、排列并测定胶原含量,冻干及金属镀膜后在扫描电子显微镜下观察真皮层胶原纤维微观形态。采用原子力显微镜在液相下测量2组瘢痕组织硬度。取2组瘢痕组织,分离和培养Fb,采用倒置相差显微镜观察其形态,并采用细胞免疫荧光法检测桩蛋白的表达以反映细胞形态,采用细胞免疫荧光法检测促纤维化蛋白α平滑肌肌动蛋白(α-SMA)、转化生长因子β1(TGF-β1)和Ⅰ型胶原表达及机械力转导相关蛋白Yes相关蛋白(YAP)和增殖相关蛋白Ki67的表达,采用实时荧光定量反转录PCR法检测促纤维化基因TGF-β1、α-SMA和Ⅰ型胶原,抑制纤维化基因TGF-β3及机械力转导相关基因Rho相关激酶1(ROCK1)和YAP mRNA表达。对数据行单因素方差分析、LSD-t检验。 结果 HE染色可见,正常皮肤表皮层凹凸不平,真皮层可见血管和汗腺等附属器;年轻组、年长组瘢痕组织表皮层均较为扁平,真皮层血管和汗腺等附属器罕见。Masson染色和扫描电子显微镜下可见,正常皮肤胶原纤维排列松散、无序,而2组瘢痕组织胶原纤维排列均较为致密、整齐,且年轻组瘢痕组织胶原纤维较年长组更为致密。年轻组、年长组瘢痕组织胶原含量明显高于正常皮肤组织(t=8.02、3.15,P<0.05或P<0.01),年长组瘢痕组织胶原含量明显低于年轻组(t=4.84,P<0.05)。年长组瘢痕组织真皮层硬度为(50.3±1.1)kPa,明显高于年轻组的(35.2±0.8)kPa(t=11.43,P<0.05)。2组瘢痕Fb在倒置相差显微镜下及经细胞免疫荧光法观察,在形态上无明显差异。年长组瘢痕Fb细胞质中Ⅰ型胶原和TGF-β1表达较年轻组明显升高,2组瘢痕Fb细胞质中α-SMA表达相近。年长组瘢痕Fb细胞质和细胞核中YAP表达较年轻组明显增多,2组瘢痕Fb细胞核中Ki67表达无明显差异。年长组瘢痕Fb中TGF-β1和Ⅰ型胶原mRNA表达量明显高于年轻组(t=2.87、4.85,P<0.05或P<0.01),TGF-β3 mRNA表达量明显低于年轻组(t=3.36,P<0.05),α-SMA mRNA表达量与年轻组无明显差异(t=1.14,P>0.05)。年长组瘢痕Fb中ROCK1和YAP mRNA表达量明显高于年轻组(t=2.98、7.60,P<0.05或P<0.01)。 结论 年长者皮肤损伤后更容易发生瘢痕愈合,其分子机制可能是由于创面愈合过程中会产生硬度较高的细胞外基质成分使得组织硬度增加,从而激活ROCK和YAP/转录共激活因子PDZ结合基序基因的表达,进而促进促纤维化基因和蛋白的表达。 Abstract:Objective To explore the effects and potential molecular mechanism of age on the stiffness and the fibrotic phenotype of fibroblasts (Fbs) of human hypertrophic scar. Methods The experimental research method was used. From January to June 2020, the surgically removed hypertrophic scar tissue of 10 scar patients (4 males and 6 females) and residual full-thickness normal skin tissue of 10 cases (5 males and 5 females, aged 7-41 years) were collected after operation in Department of Burns and Plastic Surgery of the Fourth Medical Center of the PLA General Hospital. The hypertrophic scar tissue of 6 patients aged (10.7±1.6) years was included into the young group and the hypertrophic scar tissue of 4 patients aged (40.0±2.2) years was included into the elderly group according to the age of patients. For the normal skin tissue and scar tissue in the two groups, hematoxylin eosin (HE) staining was performed to observe the tissue morphology, Masson staining was performed to observe the morphology and arrangement of collagen and quantify the content of collagen, and scanning electron microscope was used to observe the microscopic difference of dermal collagen fibers after the samples were freeze-dried and metal coated. The stiffness of scar tissue in the two groups was measured by atomic force microscope under the liquid phase. The scar tissue in the two groups was collected and the Fbs were isolated and cultured. The morphological differences of the Fbs were observed under the inverted phase contrast microscope, and the protein expression of paxillin was detected with cellular immunofluorescence to reflect the morphology of the Fbs. Cellular immunofluorescence was used to detect the expressions of pro-fibrosis protein α-smooth actin (α-SMA), transforming growth factor-β1 (TGF-β1), and type Ⅰ collagen, mechanotransduction-related protein Yes-associated protein (YAP), and the proliferation-related protein Ki67. Real-time fluorescent quantitative reverse transcription polymerase chain reaction was used to detect the mRNA expressions of pro-fibrosis genes of TGF-β1, α-SMA, and type Ⅰ collagen, fibrosis inhibiting gene of TGF-β3, and mechanotransduction-related genes of Rho-associated protein 1 (ROCK1) and YAP. Data were statistically analyzed with one-way analysis of variance and least significant difference t test. Results HE staining showed that the epidermal layer of normal skin was uneven, and blood vessels and sweat glands could be seen in the dermal layer; the epidermal layer of the scar tissue in the two groups was relatively flat, and blood vessels and sweat glands were rare. Masson staining and scanning electron microscopy showed that the collagen fibers in normal skin arranged loosely and disorderly, while the collagen fibers in scar tissue of the two groups arranged densely and orderly, and the collagen fibers in scar tissue of the young group were denser than those of the elderly group. The collagen content in scar tissue of the young group and the elderly group was significantly higher than that of the normal skin tissue (t=8.02, 3.15, P<0.05 or P<0.01), and the collagen content in scar tissue of the elderly group was significantly lower than that of the young group (t=4.84, P<0.05). The dermal stiffness of scar tissue in the elderly group was (50.3±1.1) kPa, significantly higher than (35.2±0.8) kPa in the young group (t=11.43, P<0.05). There were no obvious differences in the morphology of scar Fbs in the two groups observed under inverted phase contrast microscope and by cellular immunofluorescence. The expressions of type Ⅰ collagen and TGF-β1 in scar Fbs cytoplasm of the elderly group were significantly higher than those in the young group, while the expressions of α-SMA in scar Fbs cytoplasm were close in the two groups. The expressions of YAP in cytoplasm and nucleus of scar Fbs in the elderly group were significantly higher than those in the young group, while the expressions of Ki67 in scar Fbs nucleus of the two groups were close. The mRNA expressions of TGF-β1 and type Ⅰ collagen in scar Fbs of the elderly group were significantly higher than those in the young group (t=2.87, 4.85, P<0.05 or P<0.01), the mRNA expression of TGF-β3 in scar Fbs of the elderly group was significantly lower than that in the young group (t=3.36, P<0.05), and the mRNA expressions of α-SMA in scar Fbs of the two groups were close (t=1.14, P>0.05). The mRNA expressions of ROCK1 and YAP in scar Fbs of the elderly group were significantly higher than those in the young group (t=2.98, 7.60, P<0.05 or P<0.01). Conclusions The elderly are prone to scar healing after skin injury. The molecular mechanism may be attributed to the production of extracellular matrix components with higher stiffness, which increases tissue stiffness and thereby activates the expressions of ROCK and YAP/transcriptional co-activator with PDZ-binding motif genes, promoting pro-fibrosis gene and protein expression. -
Key words:
- Cicatrix /
- Hardness /
- Fibroblast /
- Age /
- Gene and protein of fibrosis /
- Gene and protein of mechanotransduction
-
(1)甘草酸二铵能减轻严重烫伤大鼠的肝功能损伤,并且多次使用比单次使用效果好。
(2)甘草酸二铵可能通过减轻线粒体损伤及抑制内质网应激发挥减轻肝功能损伤的作用。
烧伤是一个全世界的重大公共卫生问题,烧伤患者器官功能障碍、感染及死亡风险都明显高于其他外伤患者[1],重度烧伤患者病死率更高[2, 3]。据统计,近年来烧伤患者死亡的主要原因为MODS及脓毒血症[1],其中肝脏器官功能障碍是烧伤预后不良的早期危险因素[4]。研究表明,重度烧伤患者肝功能变化与死亡风险有明显关联,而早期采取有效措施保护肝脏及防止肝损伤进一步恶化有助于提高患者存活率[2]。烧伤后肝损伤的发生与肝脏应激性损伤、缺血缺氧及再灌注损伤、感染、肝毒性物质损伤等[5, 6, 7, 8, 9, 10, 11]息息相关,内质网应激与线粒体损伤在肝损伤的发生过程中也有着不可忽视的作用[12]。
甘草酸具有抗炎、抗肝毒性、抗病毒、抗纤维化、调节免疫等生物学特性[13, 14, 15],被证实可有效改善肝功能损伤,在临床上常被用于治疗肝炎[16]、药物性肝损伤[17]、非酒精性脂肪肝[18]等肝脏疾病。也有临床研究表明,严重烧伤患者早期使用甘草酸二铵能有效地治疗肝损伤[19, 20],但其机制并不明确。本研究团队推测,甘草酸二铵对烧伤后肝脏的保护作用可能是通过缓解内质网应激及减少线粒体损伤来实现的。本文旨在通过体内实验探讨甘草酸二铵对烫伤大鼠肝损伤的影响及其作用机制。
1. 材料与方法
本实验研究遵循国家和安徽医科大学第一附属医院有关实验动物管理和使用的规定。
1.1 动物及主要试剂与仪器来源
54只7~9周龄健康清洁级雌性SD大鼠,体重200~250 g,由安徽医科大学动物实验中心提供,许可证号:SCXK(皖)2014-003。甘草酸二铵注射液购自正大天晴药业集团股份有限公司,RIPA裂解液、二辛丁酸蛋白浓度测定试剂盒(增强型)购自上海碧云天生物技术有限公司,兔抗大鼠葡萄糖调节蛋白78(GRP78)多克隆抗体购自美国Abcam公司,兔抗大鼠转录激活因子4(ATF4)多克隆抗体、兔抗大鼠B淋巴细胞瘤-2(Bcl-2)多克隆抗体、兔抗大鼠Bcl-2相关X蛋白(Bax)多克隆抗体购自武汉三鹰生物技术有限公司,兔抗大鼠蛋白激酶R样内质网激酶(PERK)单克隆抗体购自美国Cell Signaling公司,兔源性GAPDH多克隆抗体购自北京爱必信生物技术有限公司,辣根过氧化物酶(HRP)标记的山羊抗兔IgG多克隆抗体购自北京中杉金桥生物技术有限公司,大鼠鸟氨酸氨甲酰基转移酶(OCT)ELISA检测试剂盒购自上海酶联生物科技有限公司,RNA反转录试剂盒、SYBR Green定量PCR荧光染料购自南京诺唯赞生物科技有限公司。
CKX53型倒置相差显微镜购自日本Olympus公司,PowerPac Basic型琼脂糖凝胶电泳仪购自美国伯乐公司,SMR16.1型酶标仪购自武汉优尔生生命科学装备有限公司,4500SF型凝胶数码成像分析系统购自上海Tanon科技有限公司,NANODROP 2000c型超微量蛋白核酸检测仪购自美国赛默飞世尔科技公司,AG22331型PCR扩增仪购自美国Eppendorf公司,Mx300P型实时荧光定量PCR仪购自德国罗氏科技有限公司,AU680型全自动生化检测分析仪购自贝克曼库尔特商贸(中国)有限公司。
1.2 动物模型制备、分组及标本采集
取54只大鼠,在通风、恒温、恒湿环境下适应性饲养10 d。按随机数字表法将大鼠分为假伤组、单纯烫伤组、烫伤+甘草酸二铵组,每组18只。按照3 mL/kg腹腔注射100 g/L水合氯醛将大鼠麻醉(麻醉方法下同),剃除背部毛发。单纯烫伤组及烫伤+甘草酸二铵组大鼠背部在98 ℃的水浴锅中持续浸泡12 s,造成30%TBSAⅢ度烫伤(经病理切片证实);假伤组大鼠背部在37 ℃水浴锅中持续浸泡15 s模拟致伤。假伤组大鼠伤后不进行特殊处理;单纯烫伤组及烫伤+甘草酸二铵组大鼠伤后立即按Parkland公式经腹腔注射补充乳酸林格液4 mL·kg-1·%TBSA-1抗休克,其中前8 h补充液体总量的1/2,后16 h补充剩余的1/2;烫伤+甘草酸二铵组大鼠分别于伤后1、25、49 h经腹腔注射50 mg/kg甘草酸二铵溶液。伤后24、48、72 h,每组各取6只大鼠,麻醉后取3~5 mL腹主动脉血,以2 500×g离心10 min后取上清液,于-80 ℃冰箱保存;大鼠经腹主动脉放血处死后,取肝左外叶,部分于40 g/L多聚甲醛中固定保存,部分于-80 ℃冰箱保存。
1.3 血生物化学指标检测
取3组大鼠伤后各时间点血清,于4 ℃冰箱过夜,于次日使用全自动生化检测分析仪检测血清肝功能损伤相关指标AST、ALT、乳酸脱氢酶(LDH)、总蛋白、白蛋白含量。另取3组大鼠伤后各时间点血清,37 ℃水浴融化,按ELISA检测试剂盒说明书步骤检测线粒体损伤指标OCT含量。
1.4 肝组织病理学变化观察
取储存于多聚甲醛中的3组大鼠伤后72 h肝组织,送至武汉赛维尔生物科技有限公司制作病理切片(厚度为4 μm)并进行HE染色,于400倍光学显微镜下观察肝组织病理学变化。
1.5 实时荧光定量RT-PCR法检测肝组织中内质网应激和线粒体损伤相关标志物的mRNA表达
取-80 ℃冰箱中保存的3组大鼠伤后各时间点适量肝组织,裂解后提取总RNA,并测定浓度和纯度,经反转录合成互补DNA。采用Primer3(
http://primer3.ut.ee )自行设计引物并由通用生物(安徽)股份有限公司合成,引物序列及产物大小见表1。采用实时荧光定量PCR仪检测ATF4、GRP78、PERK、Bcl-2、Bax的mRNA表达。以GAPDH为内参照,采用Δ循环阈值(Ct)法分析数据,计算各基因的相对表达量,即2-ΔΔCt。表1 大鼠肝组织中内质网应激和线粒体损伤相关标志物的引物序列及产物大小基因名称 引物序列(5'→3') 产物大小(bp) GAPDH 上游:AGACAGCCGCATCTTCTTGT 207 下游:CTTGCCGTGGGTAGAGTCAT ATF4 上游:TCCTCGATACCAGCAAATCC 64 下游:ACCCATGAGGTTTGAAGTGC GRP78 上游:GATGTAGGCACGGTGGTTGG 131 下游:GTGAAGGCCACATACGACGG PERK 上游:CCAGGCATCGTGAGGTATTT 160 下游:GATCCATCTGCCGGATCTTA Bcl-2 上游:ATACCTGGGCCACAAGTGAG 214 下游:TGATTTGACCATTTGCCTGA Bax 上游:CGAGCTGATCAGAACCATCA 191 下游:CTCAGCCCATCTTCTTCCAG 注:GAPDH 为3-磷酸甘油醛脱氢酶,ATF4为转录激活因子4,GRP78 为葡萄糖调节蛋白78,PERK为蛋白激酶R样内质网激酶,Bcl-2为B淋巴细胞瘤-2,Bax为Bcl-2相关X蛋白 1.6 蛋白质印迹法检测肝组织中内质网应激和线粒体损伤相关标志物的蛋白表达
取适量冻存于-80 ℃冰箱的假伤组大鼠伤后72 h和烫伤2组大鼠伤后各时间点肝组织,提取组织蛋白,并测定蛋白浓度,进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳,采用湿法转膜,用50 g/L脱脂牛奶室温封闭1~2 h。加入一抗兔抗大鼠GRP78多克隆抗体(稀释比为1∶1 000)、兔抗大鼠ATF4多克隆抗体(稀释比为1∶1 000)、兔抗大鼠Bcl-2多克隆抗体(稀释比为1∶1 000)、兔抗大鼠Bax多克隆抗体(稀释比为1∶6 000)、兔抗大鼠PERK单克隆抗体(稀释比为1∶1 000)和兔源性GAPDH多克隆抗体(稀释比为1∶3 000),于4 ℃条件下孵育过夜。洗涤后加入HRP标记的山羊抗兔 IgG多克隆二抗(稀释比为 1∶10 000),室温孵育1.5 h。洗涤后行化学发光显影,采用Image J 图像分析软件(美国国立卫生研究院)行灰度分析,以GAPDH为内参照,观察GRP78、ATF4、PERK、Bcl-2、Bax蛋白表达。
1.7 统计学处理
采用SPSS 26.0统计软件进行分析。计量资料数据均符合正态分布,以
2. 结果
2.1 血清肝功能损伤相关指标的含量
与假伤组比较,单纯烫伤组大鼠伤后各时间点血清中AST、ALT、LDH含量均明显升高(P<0.01),总蛋白、白蛋白含量均明显降低(P<0.05或P<0.01)。伤后24 h,相较于单纯烫伤组,烫伤+甘草酸二铵组大鼠血清中AST含量明显下降(P<0.05),而ALT、LDH、总蛋白和白蛋白含量均无明显变化(P>0.05)。伤后48 h,相较于单纯烫伤组,烫伤+甘草酸二铵组大鼠血清中AST、ALT、LDH含量均明显下降(P<0.01),总蛋白含量明显升高(P<0.01),而白蛋白含量无明显变化(P>0.05)。伤后72 h,相较于单纯烫伤组,烫伤+甘草酸二铵组大鼠血清中AST、ALT、LDH含量均明显下降(P<0.01),总蛋白和白蛋白含量均明显升高(P<0.01)。见表2。
表2 假伤组与严重烫伤2组大鼠伤后各时间点肝功能损伤相关指标含量比较(U/L,组别与时间点 鼠数(只) AST ALT LDH 总蛋白 白蛋白 假伤组 18 24 h 60±5 23±3 126±40 53.3±1.0 31.6±1.0 48 h 65±11 25±4 129±41 51.4±1.5 30.0±1.3 72 h 73±15 27±6 124±39 48.8±2.6 30.5±1.1 单纯烫伤组 18 24 h 1 520±291a 285±48a 2 257±568a 48.9±2.9d 22.7±1.6a 48 h 928±91a 211±23a 687±312a 43.3±2.0a 22.7±1.4a 72 h 515±125a 127±30a 568±208a 34.4±7.1a 18.2±3.2a 烫伤+甘草酸二铵组 18 24 h 1 228±63b 248±40 1 817±422 50.1±3.2 24.1±3.0 48 h 628±41c 136±10c 269±59c 47.5±2.5c 24.1±1.6 72 h 186±27c 64±16c 145±36c 46.0±2.4c 24.1±2.5c F 1值 121.08 92.62 45.38 4.74 33.72 P 1值 <0.001 <0.001 <0.001 0.025 <0.001 F 2值 344.11 236.87 14.83 24.29 42.84 P 2值 <0.001 <0.001 <0.001 <0.001 <0.001 F 3值 57.07 37.41 24.42 16.74 37.99 P 3值 <0.001 <0.001 <0.001 <0.001 <0.001 注:各组各时间点鼠数均为6只;AST为天冬氨酸转氨酶,ALT为丙氨酸转氨酶,LDH为乳酸脱氢酶;AST、ALT、LDH、总蛋白、白蛋白含量处理因素主效应,F值分别为309.40、242.67、68.65、35.33、105.21,P值均<0.001;时间因素主效应,F值分别为162.92、88.07、97.16、25.46、4.06,P值分别为<0.001、<0.001、<0.001、<0.001、0.024;两者交互作用,F值分别为42.25、24.72、24.30、5.21、3.46,P值分别为<0.001、<0.001、<0.001、0.002、0.015;F 1值、P 1值,F 2值、P 2值,F 3值、P 3值分别为3组大鼠伤后24、48、72 h各指标总体比较所得;与假伤组比较,a P<0.01,d P<0.05;与单纯烫伤组比较,b P<0.05,c P<0.01 2.2 血清OCT含量
与假伤组比较,单纯烫伤组大鼠伤后各时间点血清OCT含量均明显升高(P<0.01);与单纯烫伤组相比,烫伤+甘草酸二铵组大鼠伤后各时间点血清中OCT的含量均明显降低(P<0.05或P<0.01)。见表3。
表3 假伤组与严重烫伤2组大鼠伤后各时间点血清OCT含量比较(U/L,组别 鼠数(只) 24 h 48 h 72 h 假伤组 18 15.1±2.5 15.7±2.6 16.4±3.7 单纯烫伤组 18 48.5±3.9a 40.8±2.4a 38.7±2.0a 烫伤+甘草酸二铵组 18 39.0±4.5b 31.8±2.0c 22.1±2.6c F值 127.48 179.10 127.42 P值 <0.001 <0.001 <0.001 注:各组各时间点鼠数均为6只;OCT为鸟氨酸氨甲酰基转移酶;处理因素主效应,F=355.49,P<0.001;时间因素主效应,F=35.30,P<0.001;两者交互作用,F=20.37,P<0.001;F值、P值为3组伤后各时间点OCT含量总体比较所得;与假伤组比较,a P<0.01;与单纯烫伤组比较,b P<0.05,c P<0.01 2.3 肝组织病理学变化
伤后72 h,假伤组大鼠肝组织中细胞形态正常,排列规则,未见明显炎症细胞浸润;单纯烫伤组大鼠肝组织中细胞排列紊乱,可见明显胞质疏松,并伴有弥漫性的脂肪病变和中等量炎症细胞浸润;烫伤+甘草酸二铵组大鼠肝组织中细胞排列较规则,可见散在的脂肪变性,伴有少量散在炎症细胞浸润。见图1。
 1 假伤组与严重烫伤2组大鼠伤后72 h肝组织病理学变化 苏木精-伊红×400。1A.假伤组肝细胞呈放射状排列,排列规则,未见明显炎症细胞浸润;1B.单纯烫伤组肝细胞排列紊乱,胞质疏松,伴有中等量炎症细胞浸润;1C.烫伤+甘草酸二铵组肝细胞排列较规则,有少量炎症细胞浸润
1 假伤组与严重烫伤2组大鼠伤后72 h肝组织病理学变化 苏木精-伊红×400。1A.假伤组肝细胞呈放射状排列,排列规则,未见明显炎症细胞浸润;1B.单纯烫伤组肝细胞排列紊乱,胞质疏松,伴有中等量炎症细胞浸润;1C.烫伤+甘草酸二铵组肝细胞排列较规则,有少量炎症细胞浸润2.4 肝组织中线粒体损伤相关标志物的mRNA和蛋白表达
与假伤组比较,单纯烫伤组大鼠伤后24、48、72 h肝组织Bcl-2 mRNA(P<0.05或P<0.01)和蛋白表达均明显减少,Bax的mRNA(P<0.01)和蛋白表达均明显增加。与单纯烫伤组比较,烫伤+甘草酸二铵组大鼠伤后24 h肝组织Bcl-2、Bax的mRNA(P>0.05)和蛋白表达均无明显变化;烫伤+甘草酸二铵组大鼠伤后48 h肝组织Bax 的mRNA(P<0.05)和蛋白表达均明显减少,Bcl-2的mRNA(P>0.05)和蛋白表达均无明显变化;烫伤+甘草酸二铵组大鼠伤后72 h肝组织Bax的mRNA(P<0.01)和蛋白表达均明显减少,Bcl-2的mRNA(P<0.01)和蛋白表达均明显增加。见图2。
 2 假伤组与烫伤2组大鼠伤后各时间点肝组织中线粒体损伤相关标志物的mRNA和蛋白表达比较。2A.Bcl-2的mRNA表达(样本数均为6,
2 假伤组与烫伤2组大鼠伤后各时间点肝组织中线粒体损伤相关标志物的mRNA和蛋白表达比较。2A.Bcl-2的mRNA表达(样本数均为6,2.5 肝组织中内质网应激相关标志物的mRNA和蛋白表达
与假伤组比较,单纯烫伤组大鼠伤后各时间点肝组织ATF4、GRP78、PERK的mRNA(P<0.05或P<0.01)和蛋白表达均明显增加。与单纯烫伤组相比,烫伤+甘草酸二铵组大鼠伤后24 h肝组织ATF4、GRP78、PERK的mRNA(P>0.05)和蛋白表达均无明显变化;烫伤+甘草酸二铵组大鼠伤后48 h肝组织ATF4的mRNA(P<0.01)和蛋白表达均明显减少,但GRP78、PERK的mRNA(P>0.05)和蛋白表达均无明显变化;烫伤+甘草酸二铵组大鼠伤后72 h肝组织ATF4、GRP78、PERK的mRNA(P<0.05或P<0.01)和蛋白表达均明显减少。见图3。
 3 假伤组与烫伤2组大鼠伤后各时间点肝组织中内质网应激相关标志物的mRNA和蛋白表达比较。3A.PERK的mRNA表达(样本数均为6,
3 假伤组与烫伤2组大鼠伤后各时间点肝组织中内质网应激相关标志物的mRNA和蛋白表达比较。3A.PERK的mRNA表达(样本数均为6,3. 讨论
肝脏在烧伤后的机体代谢、炎症反应及免疫调节过程中发挥着巨大作用,肝损伤是重度烧伤患者常见的并发症之一[21]。甘草作为中草药,一直被广泛用于治疗肝脏相关疾病,而甘草酸是甘草中最为重要的生物活性化合物[22, 23]。甘草酸二铵作为第3代甘草酸制剂,是18β-甘草酸与18α-甘草酸的混合构型体,具有水溶性强、扩散性强等特点。有研究证实,甘草酸二铵能有效减轻由刀豆蛋白A诱导的小鼠肝损伤[23]。还有临床研究表明,严重烧伤患者使用甘草酸二铵能显著减轻肝损伤[19, 20]。本研究建立了30%TBSA的Ⅲ度烫伤SD大鼠模型,并观察了腹腔注射甘草酸二铵对大鼠烫伤后肝损伤的影响。结果显示,单纯烫伤组大鼠伤后各时间点血清中AST、ALT、LDH等肝功能损伤指标的含量均较假伤组显著升高,血清总蛋白、白蛋白含量均较假伤组明显减少,HE染色显示单纯烫伤组大鼠肝脏出现脂肪病变、炎症反应,表明严重烧伤大鼠伤后存在肝损伤。
白蛋白是由肝实质细胞合成,是血浆中含量最多的蛋白,肝脏受损后白蛋白及总蛋白的合成将会减少,因此血清中白蛋白及总蛋白含量减少提示着肝功能受损;但对严重烧伤患者来说,血清白蛋白及总蛋白的减少主要是由创面渗出及低血容量性休克所致,因此这2项指标减少只能在一定程度上提示肝功能受损,需结合其他指标一起观察。本研究结果表明,当注射1次甘草酸二铵后,大鼠血清中总蛋白及白蛋白含量均无明显改变,但当连续注射3次甘草酸二铵后,大鼠血清中总蛋白及白蛋白含量均明显升高,提示连续多次使用甘草酸二铵可减轻大鼠肝功能损伤。
线粒体是真核细胞中一个极为重要的细胞器。在维持细胞生理功能的过程中,线粒体结构和功能的完整性起着尤为重要的作用[24],它不仅能合成ATP为细胞供能,还在细胞损伤过程中起着关键作用[25]。OCT是一种主要存在于肝细胞线粒体内的标志性酶,肝损伤后肝细胞膜及线粒体外膜通透性增加,OCT随之释放入血,因此血清内OCT含量增加提示肝细胞线粒体受损[11,23]。本研究结果显示,单纯烫伤组大鼠伤后各时间点血清OCT含量均较假伤组显著升高,说明严重烫伤大鼠伤后存在线粒体损伤,而注射甘草酸二铵后大鼠血清OCT含量均明显降低,与相关研究结果[12,26]一致。且连续注射甘草酸二铵3次后,大鼠血清中OCT含量下降得更为明显,这说明多次使用甘草酸二铵对线粒体损伤的改善作用较单次注射更为明显。本研究团队还检测了肝组织中Bcl-2和Bax的表达水平,Bcl-2及Bax是Bcl-2蛋白家族中的重要成员[25]。Bcl-2蛋白家族定位在线粒体外膜上,其促凋亡和抗凋亡成员在线粒体表面相互作用。当细胞内出现应激信号时,促凋亡蛋白如胞质Bax蛋白被激活并转移到线粒体表面,在线粒体表面中和抗凋亡蛋白如Bcl-2膜蛋白,从而触发线粒体膜通透性增加,继而诱导凋亡[25]。本研究结果显示,单纯烫伤组大鼠伤后各时间点Bcl-2 mRNA和蛋白表达均较假伤组减少,而Bax mRNA和蛋白表达均较假伤组增加;注射1次甘草酸二铵后,大鼠肝组织中Bcl-2及Bax的mRNA及蛋白表达均无明显改变,但连续注射3次甘草酸二铵后,大鼠肝组织中Bcl-2的mRNA及蛋白表达均明显增加,Bax的mRNA及蛋白表达均明显减少。以上结果提示连续多次使用甘草酸二铵可减轻烫伤引起的线粒体损伤。
研究表明,严重烧伤会导致肝脏结构和功能完整性破坏,钙稳态失衡,肝凋亡和内质网稳态失衡等改变[27],继而使肝脏功能出现紊乱,其中内质网过度应激和线粒体损伤是2种重要的通路。内质网是细胞中不可或缺的亚细胞单位,负责蛋白质的合成、修饰、折叠、组装和运输[28, 29]。当在炎症、氧化应激和缺氧等压力条件下,错误折叠或未折叠蛋白不断累积,内质网稳态被打破,功能出现紊乱,即为内质网应激[30]。细胞为缓解内质网应激,帮助内质网恢复功能,会触发未折叠蛋白反应(UPR)。本研究主要关注的是UPR中的PERK通路。在正常情况下,PERK与GRP78紧密连接,保持在非活力状态;当内质网稳态受到干扰时,PERK会与GRP78解离,磷酸化后形成二聚体,即磷酸化PERK(p-PERK)。 p-PERK将下游的真核起始因子2α磷酸化,促进ATF4翻译,从而启动一系列基因转录来缓解内质网压力[31]。然而,内质网应激过于严重时,ATF4会诱导促细胞凋亡程序的运行[30]。有研究表明,严重烧伤可激活内质网应激信号转导途径 [23]。同样,本研究结果显示,严重烫伤大鼠肝组织中内质网应激相关标志物GRP78、ATF4、PERK的mRNA和蛋白表达水平在烫伤后显著升高;注射1次甘草酸二铵后,肝组织中ATF4、GRP78及PERK的mRNA及蛋白表达均无明显改变;连续注射2次甘草酸二铵后,肝组织中ATF4的mRNA及蛋白表达有所下降,但GRP78及PERK的mRNA及蛋白表达均无明显改变;连续注射3次甘草酸二铵后,肝组织中ATF4、GRP78及PERK的mRNA及蛋白表达均明显降低。以上结果提示连续多次使用甘草酸二铵可减轻烫伤引起的内质网应激。
综上所述,严重烫伤会起内质网应激与线粒体损伤,造成肝功能损伤;甘草酸二铵可明显减弱肝内质网应激,同时改善线粒体功能,继而有效降低严重烫伤后大鼠肝损伤的程度。然而,目前尚不清楚线粒体损伤是否诱导内质网应激,或者内质网应激是否诱导线粒体损伤。有研究显示,线粒体损伤和内质网应激可能是相互联系的[32],而甘草酸二铵具有改善肝线粒体功能的作用并减轻肝内质网应激,从而减轻肝损伤,但其潜在关联仍需进一步实验深入研究。
所有作者均声明不存在利益冲突 -
参考文献
(36) [1] KirkwoodTB. Understanding the odd science of aging[J]. Cell, 2005,120(4):437-447.DOI: 10.1016/j.cell.2005.01.027. [2] BakerDJ, PetersenRC. Cellular senescence in brain aging and neurodegenerative diseases: evidence and perspectives[J]. J Clin Invest, 2018,128(4):1208-1216. DOI: 10.1172/JCI95145. [3] RamirezT, LiYM, YinS, et al. Aging aggravates alcoholic liver injury and fibrosis in mice by downregulating sirtuin 1 expression[J]. J Hepatol, 2017,66(3):601-609. DOI: 10.1016/j.jhep.2016.11.004. [4] ShusterS, BlackMM, Mc VitieE. The influence of age and sex on skin thickness, skin collagen and density[J]. Br J Dermatol, 1975,93(6):639-643. DOI: 10.1111/j.1365-2133.1975.tb05113.x. [5] MooreAL, MarshallCD, BarnesLA, et al. Scarless wound healing: transitioning from fetal research to regenerative healing[J]. Wiley interdiscip Rev Dev Biol, 2018,7(2): 10.1002/wdev.309. DOI: 10.1002/wdev.309. [6] WillyardC. Unlocking the secrets of scar-free skin healing[J]. Nature, 2018,563(7732):S86-88. DOI: 10.1038/d41586-018-07430-w. [7] SolonJ, LeventalI, SenguptaK, et al. Fibroblast adaptation and stiffness matching to soft elastic substrates[J]. Biophys J, 2007,93(12):4453-4461. DOI: 10.1529/biophysj.106.101386. [8] WellsRG. Tissue mechanics and fibrosis[J]. Biochim Biophys Acta, 2013,1832(7):884-890. DOI: 10.1016/j.bbadis.2013.02.007. [9] DasguptaI, McCollumD. Control of cellular responses to mechanical cues through YAP/TAZ regulation[J]. J Biol Chem, 2019,294(46):17693-17706. DOI: 10.1074/jbc.REV119.007963. [10] DupontS. Role of YAP/TAZ in cell-matrix adhesion-mediated signalling and mechanotransduction[J]. Exp Cell Res, 2016,343(1):42-53. DOI: 10.1016/j.yexcr.2015.10.034. [11] LichtmanMK, Otero-VinasM, FalangaV. Transforming growth factor beta (TGF-β) isoforms in wound healing and fibrosis[J]. Wound Repair Regen, 2016,24(2):215-222. DOI: 10.1111/wrr.12398. [12] LiuF, LagaresD, ChoiKM, et al. Mechanosignaling through YAP and TAZ drives fibroblast activation and fibrosis[J]. Am J Physiol Lung Cell Mol Physiol, 2015,308(4):L344-357. DOI: 10.1152/ajplung.00300.2014. [13] SzetoSG, NarimatsuM, LuM, et al. YAP/TAZ are mechano- regulators of TGF-β-Smad signaling and renal fibrogenesis[J]. J Am Soc Nephrol, 2016,27(10):3117-3128.DOI: 10.1681/asn.2015050499. [14] AbramczykH, ImielaA, Brozek-PluskaB, et al. Advances in Raman imaging combined with AFM and fluorescence microscopy are beneficial for oncology and cancer research[J]. Nanomedicine (Lond), 2019,14(14):1873-1888. DOI: 10.2217/nnm-2018-0335. [15] BhushanB. Nanotribological and nanomechanical properties of skin with and without cream treatment using atomic force microscopy and nanoindentation[J]. J Colloid Interface Sci, 2012,367(1):1-33. DOI: 10.1016/j.jcis.2011.10.019. [16] StylianouA, LekkaM, StylianopoulosT. AFM assessing of nanomechanical fingerprints for cancer early diagnosis and classification: from single cell to tissue level[J]. Nanoscale, 2018,10(45):20930-20945. DOI: 10.1039/c8nr06146g. [17] GauglitzGG, KortingHC, PavicicT, et al. Hypertrophic scarring and keloids: pathomechanisms and current and emerging treatment strategies[J]. Mol Med, 2011,17(1/2):113-125. DOI: 10.2119/molmed.2009.00153. [18] PuglieseE, CoentroJQ, RaghunathM, et al. Wound healing and scar wars[J]. Adv Drug Deliv Rev, 2018,129:1-3. DOI: 10.1016/j.addr.2018.05.010. [19] 张波, 王正国, 朱佩芳. 皮肤无瘢痕愈合机制的研究进展[J]. 中华烧伤杂志, 2002,18(5):318-320. [20] KöseO, WaseemA. Keloids and hypertrophic scars: are they two different sides of the same coin?[J]. Dermatol Surg, 2008,34(3):336-346. DOI: 10.1111/j.1524-4725.2007.34067.x. [21] RittiéL, FisherGJ. Natural and sun-induced aging of human skin[J]. Cold Spring Harb Perspect Med, 2015,5(1):a015370. DOI: 10.1101/cshperspect.a015370. [22] GeorgesPC, HuiJJ, GombosZ, et al. Increased stiffness of the rat liver precedes matrix deposition: implications for fibrosis[J]. Am J Physiol Gastrointest Liver Physiol, 2007,293(6):G1147-1154. DOI: 10.1152/ajpgi.00032.2007. [23] Viji BabuPK, RiannaC, BelgeG, et al. Mechanical and migratory properties of normal, scar, and Dupuytren's fibroblasts[J]. J Mol Recognit, 2018,31(9):e2719. DOI: 10.1002/jmr.2719. [24] TschumperlinDJ, LagaresD. Mechano-therapeutics: targeting mechanical signaling in fibrosis and tumor stroma[J]. Pharmacol Ther, 2020,212:107575. DOI: 10.1016/j.pharmthera.2020.107575. [25] TschumperlinDJ, LigrestiG, HilscherMB, et al. Mechanosensing and fibrosis[J]. J Clin Invest, 2018,128(1):74-84. DOI: 10.1172/JCI93561. [26] WuH, YuY, HuangH, et al. Progressive pulmonary fibrosis is caused by elevated mechanical tension on alveolar stem cells[J]. Cell, 2020,180(1):107-121.e17. DOI: 10.1016/j.cell.2019.11.027. [27] YaoB, ZhuDZ, CuiXL, et al. Modeling human hypertrophic scars with 3D preformed cellular aggregates bioprinting[J]. Bioactive Materials, 2021,In press. DOI: 10.1016/j.bioactmat.2021.09.004. [28] HinzB. Myofibroblasts[J]. Exp Eye Res, 2016,142:56-70. DOI: 10.1016/j.exer.2015.07.009. [29] HinzB, McCullochCA, CoelhoNM. Mechanical regulation of myofibroblast phenoconversion and collagen contraction[J]. Exp Cell Res, 2019,379(1):119-128.DOI: 10.1016/j.yexcr.2019.03.027. [30] KnipeRS, ProbstCK, LagaresD, et al. The Rho kinaseis oforms ROCK1 and ROCK2 each contribute to the development of experimental pulmonary fibrosis[J]. Am J Respir Cell Mol Biol, 2018,58(4):471-481. DOI: 10.1165/rcmb.2017-0075OC. [31] ShiJ, SurmaM, YangY, et al. Disruption of both ROCK1 and ROCK2 genes in cardiomyocytes promotes autophagy and reduces cardiac fibrosis during aging[J]. FASEB J, 2019,33(6):7348-7362. DOI: 10.1096/fj.201802510R. [32] ZhouY, HuangX, HeckerL, et al. Inhibition of mechanosensitive signaling in myofibroblasts ameliorates experimental pulmonary fibrosis[J]. J Clin Invest, 2013,123(3):1096-1108. DOI: 10.1172/JCI66700. [33] NoguchiS, SaitoA, NagaseT. YAP/TAZ signaling as a molecular link between fibrosis and cancer[J]. Int J Mol Sci, 2018,19(11):3674. DOI: 10.3390/ijms19113674. [34] Picollet-D'hahanN, DolegaME, LiguoriL, et al. A 3D toolbox to enhance physiological relevance of human tissue models[J]. Trends Biotechnol, 2016,34(9):757-769.DOI: 10.1016/j.tibtech.2016.06.012. [35] YanWC, DavoodiP, VijayavenkataramanS, et al. 3D bioprinting of skin tissue: from pre-processing to final product evaluation[J]. Adv Drug Deliv Rev, 2018,132:270-295.DOI: 10.1016/j.addr.2018.07.016. [36] 朱冬振, 王一惠, 王睿, 等. 外源性肿瘤坏死因子α对三维环境下小鼠间充质干细胞向汗腺细胞分化的影响及机制[J]. 中华烧伤杂志, 2020,36(3):187-194. DOI: 10.3760/cma.j.cn501120-20200105-00005. -

1 植皮术患者正常皮肤和2组增生性瘢痕患者瘢痕组织形态 苏木精-伊红×100,图中标尺为500 μm。1A.正常皮肤组织褶皱较为明显,表皮基底层凹凸不平,真皮层可见血管和汗腺等附属器;1B.年轻组瘢痕组织表皮层扁平,表皮基底层正常棘突消失,真皮层血管和汗腺等附属器罕见;1C.年长组瘢痕组织结构与图1B相似
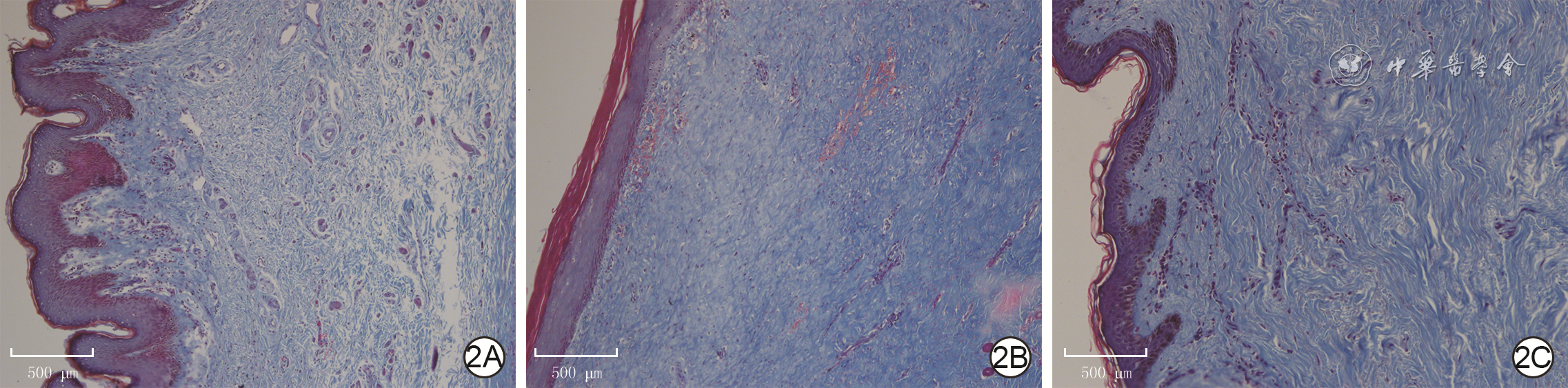
2 植皮术正常皮肤和2组增生性瘢痕患者瘢痕组织真皮层胶原排列和分布 Masson×100,图中标尺为500 μm。2A.正常皮肤组织胶原排列松散、无序;2B.年轻组瘢痕组织胶原排列致密且有序;2C.年长组瘢痕组织皮肤胶原排列有序,但较图2B稀疏且胶原含量减少

4 植皮术患者正常皮肤和2组增生性瘢痕患者瘢痕组织真皮层胶原纤维微观形态 扫描电子显微镜×500,图中标尺为200 μm。4A.正常皮肤组织胶原纤维排列松散、无序并相互交织;4B.年轻组瘢痕组织真皮层胶原纤维致密且排列有序;4C.年长组瘢痕组织皮肤胶原纤维聚集成束,且排列有序,但较图4B稀疏
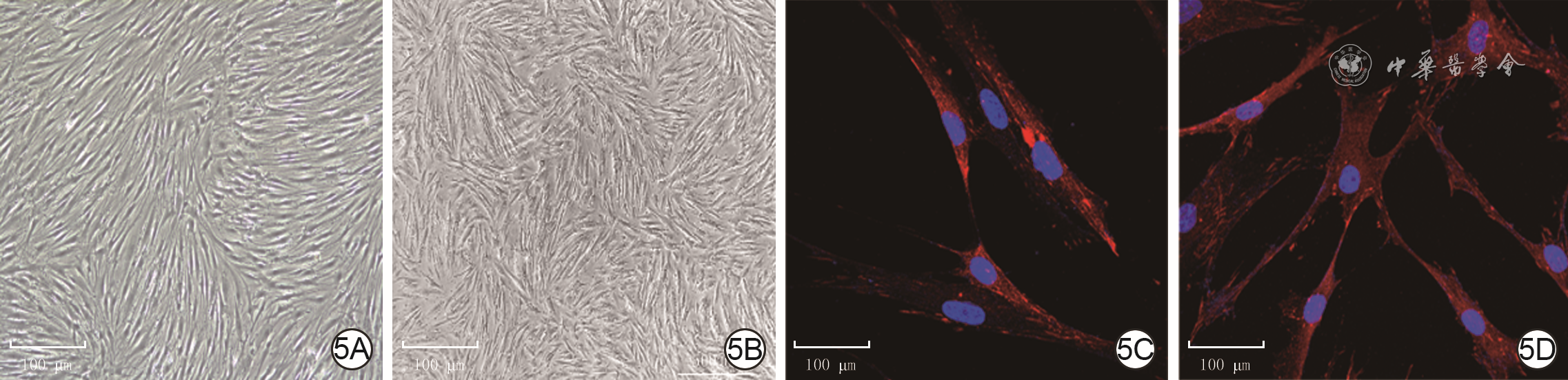
5 2组人瘢痕成纤维细胞形态学观察。5A、5B.分别为年轻组和年长组细胞形态,2组细胞均呈梭形并聚集呈巢状 倒置相差显微镜×50,图中标尺为500 μm;5C、5D.分别为年轻组和年长组细胞形态,2组细胞均呈多边形,相互重叠,并向周围伸展,细胞核呈圆形或椭圆形 Alexa Fluor 594-4',6-二脒基-2-苯基吲哚×400,图中标尺为100 μm
注:细胞核阳性染色为蓝色,桩蛋白阳性染色为红色
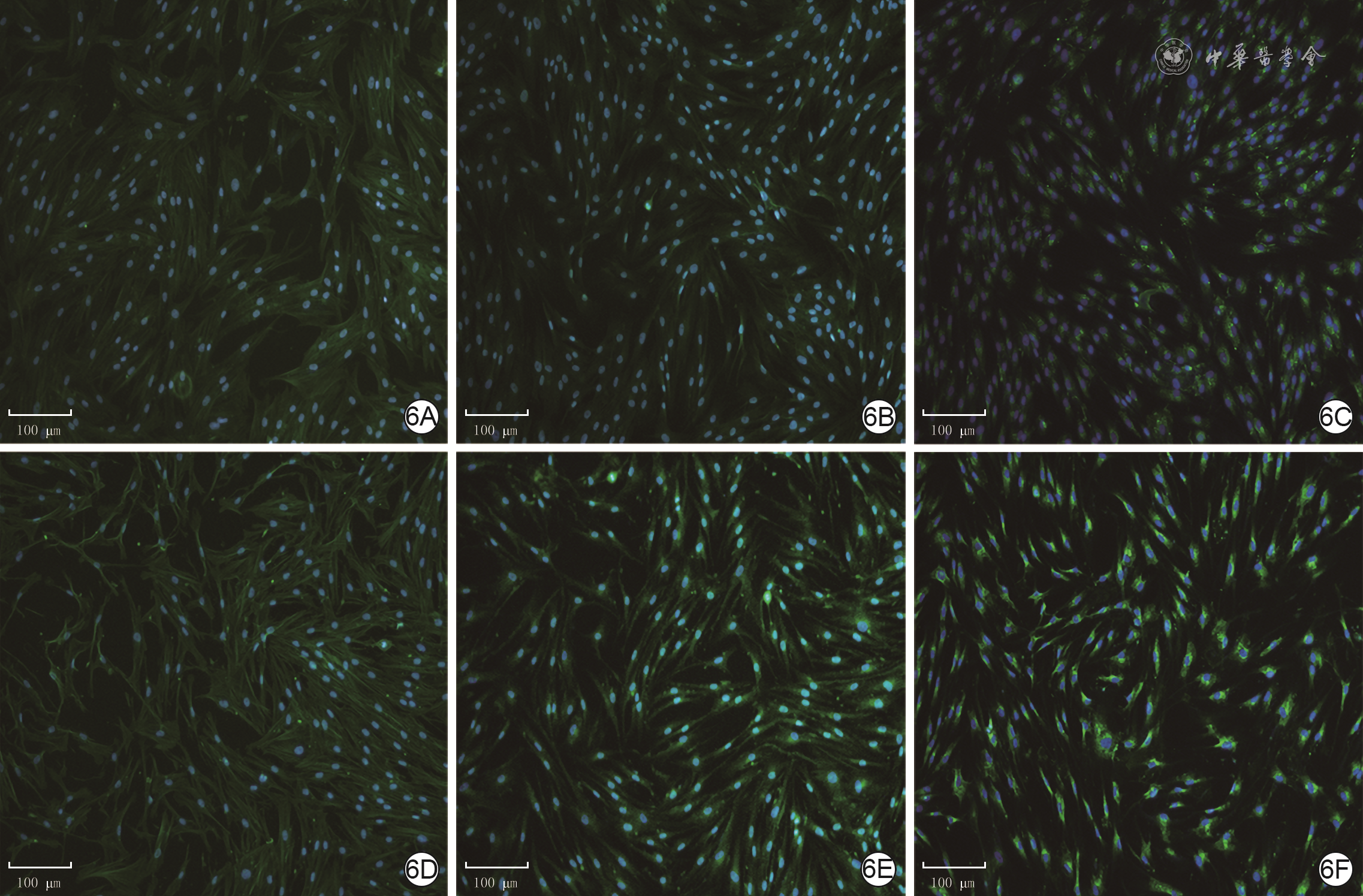
6 2组人瘢痕成纤维细胞中α-SMA、Ⅰ型胶原和TGF-β1蛋白的表达 Alexa Fluor 488-4',6-二脒基-2-苯基吲哚×100,图中标尺为100 μm。6A、6B、6C.分别为年轻组细胞质中α-SMA、Ⅰ型胶原和TGF-β1的表达;6D、6E、6F.分别为年长组细胞质中α-SMA、Ⅰ型胶原和TGF-β1的表达,α-SMA的表达与图6A相近,Ⅰ型胶原的表达较图6B明显增加,TGF-β1的表达较图6C明显增加
注:细胞核阳性染色为蓝色,α平滑肌肌动蛋白(α-SMA)、Ⅰ型胶原和转化生长因子β1(TGF-β1)阳性染色均为绿色
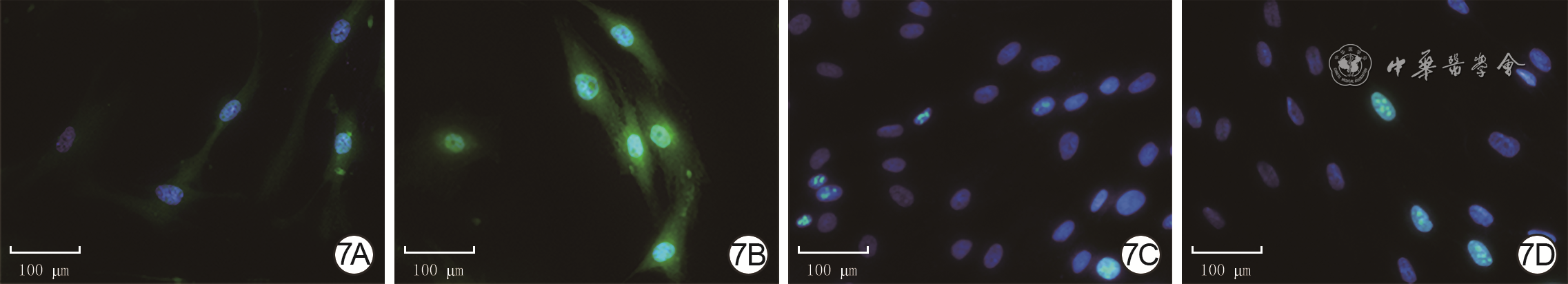
7 2组人瘢痕成纤维细胞中YAP和Ki67蛋白的表达 Alexa Fluor 488-4',6-二脒基-2-苯基吲哚×400,图中标尺为100 μm。7A、7B.分别为年轻组和年长组,YAP蛋白广泛分布于细胞质和细胞核,且图7B中YAP蛋白较图7A增加;7C、7D.分别为年轻组和年长组,Ki67蛋白分布于细胞核,且图7D中Ki67蛋白表达与图7C无明显差异
注:细胞核阳性染色为蓝色,Yes相关蛋白(YAP)和Ki67阳性染色均为绿色

8 实时荧光定量反转录PCR法检测2组人瘢痕成纤维细胞中纤维化相关基因和机械力转导相关基因的表达(
注:TGF为转化生长因子,α-SMA为α平滑肌肌动蛋白,ROCK1为Rho相关激酶1,YAP为Yes相关蛋白;与年轻组比较,aP<0.05,bP<0.01
表1 实时荧光定量反转录PCR检测2组患者瘢痕成纤维细胞中纤维化相关基因和机械力转导相关基因的表达
基因名称 引物序列(5’→3’) 产物大小(bp) GAPDH 上游:GGAGCGAGATCCCTCCAAAAT 197 下游:GGCTGTTGTCATACTTCTCATGG TGF-β1 上游:CAATTCCTGGCGATACCTCAG 86 下游:GCACAACTCCGGTGACATCAA TGF-β3 上游:ACTTGCACCACCTTGGACTTC 114 下游:GGTCATCACCGTTGGCTCA α-SMA 上游:GTGTTGCCCCTGAAGAGCAT 116 下游:GCTGGGACATTGAAAGTCTCA Ⅰ型胶原 上游:GAGGGCCAAGACGAAGACATC 140 下游:CAGATCACGTCATCGCACAAC YAP 上游:TAGCCCTGCGTAGCCAGTTA 177 下游:TCATGCTTAGTCCACTGTCTGT ROCK1 上游:AACATGCTGCTGGATAAATCTGG 93 下游:TGTATCACATCGTACCATGCCT 注:GAPDH为3-磷酸甘油醛脱氢酶,TGF为转化生长因子,α-SMA为α平滑肌肌动蛋白,YAP为Yes相关蛋白,ROCK1为Rho相关激酶1  下载: 导出CSV
下载: 导出CSV
-


 下载:
下载:
计量
- 文章访问数: 385
- HTML全文浏览量: 190
- PDF下载量: 58
- 被引次数: 0
