



Regulatory effects of bio-intensity electric field on transformation of human skin fibroblasts
-
摘要:
目的 探讨生物强度电场对人皮肤成纤维细胞(HSF)转化的调节作用。 方法 采用实验研究方法。取HSF,分为经200 mV/mm电场处理6 h的200 mV/mm电场组和置于电场装置中不通电处理6 h的模拟电场组,在活细胞工作站中观察细胞形态和排列变化;记录处理0、6 h细胞数,并计算细胞数变化率;观察并计算3 h内细胞运动方向、位移速度、轨迹速度(以上实验模拟电场组样本数为34、200 mV/mm电场组样本数为30);采用免疫荧光法检测处理3 h细胞α平滑肌肌动蛋白(α-SMA)的蛋白表达(样本数为3)。取HSF分为置于电场装置中不通电处理3 h的模拟电场组和经相应强度电场处理3 h的100 mV/mm电场组、200 mV/mm电场组、400 mV/mm电场组,另取HSF分为置于电场装置中不通电处理6 h的模拟电场组和经200 mV/mm电场处理相应时间的电场处理1 h组、电场处理3 h组、电场处理6 h组,采用蛋白质印迹法检测α-SMA、增殖细胞核抗原(PCNA)的蛋白表达(样本数为3)。对数据行Mann-Whitney U检验、单因素方差分析、独立样本t检验及LSD检验。 结果 处理6 h,与模拟电场组相比,200 mV/mm电场组细胞形态拉长,并产生局部粘连;模拟电场组细胞任意排列,200 mV/mm电场组细胞呈有规律的纵向排列;2组细胞数变化率相近(P>0.05)。处理3 h内,200 mV/mm电场组细胞有明显的向正极运动趋势,模拟电场组细胞绕原点运动;与模拟电场组比较,200 mV/mm电场组细胞位移速度和轨迹速度均明显加快(Z值分别为-5.33、-5.41,P<0.01),方向性显著增强(Z=-4.39,P<0.01)。处理3 h,200 mV/mm电场组细胞α-SMA蛋白表达较模拟电场组明显增加(t=-9.81,P<0.01)。处理3 h,100 mV/mm电场组、200 mV/mm电场组、400 mV/mm电场组细胞α-SMA蛋白表达分别为1.195±0.057、1.606±0.041、1.616±0.039,均明显多于模拟电场组的0.649±0.028(P<0.01)。与100 mV/mm电场组比较,200 mV/mm电场组、400 mV/mm电场组细胞α-SMA蛋白表达均明显增加(P<0.01)。电场处理1 h组、电场处理3 h组、电场处理6 h组细胞α-SMA蛋白表达分别为0.730±0.032、1.561±0.031、1.553±0.045,均明显多于模拟电场组的0.464±0.020(P<0.01);与电场处理1 h组比较,电场处理3 h组、电场处理6 h组细胞α-SMA蛋白表达均明显增加(P<0.01)。处理3 h,与模拟电场组比较,100 mV/mm电场组、200 mV/mm电场组、400 mV/mm电场组细胞PCNA蛋白表达均明显减少(P<0.05或P<0.01);与100 mV/mm电场组比较,200 mV/mm电场组、400 mV/mm电场组细胞PCNA蛋白表达均明显减少(P<0.05或P<0.01);与200 mV/mm电场组比较,400 mV/mm电场组细胞PCNA蛋白表达明显减少(P<0.01)。与模拟电场组比较,电场处理1 h组、电场处理3 h组、电场处理6 h组细胞PCNA蛋白表达均明显减少(P<0.01);与电场处理1 h组比较,电场处理3 h组、电场处理6 h组细胞PCNA蛋白表达均明显减少(P<0.05或P<0.01);与电场处理3 h组比较,电场处理6 h组细胞PCNA蛋白表达明显减少(P<0.01)。 结论 生物强度电场可诱导HSF迁移、促进Fb向肌Fb转化,且转化有一定的时间及电场强度依赖性。 Abstract:Objective To investigate the regulatory effects of bio-intensity electric field on the transformation of human skin fibroblasts (HSFs). Methods The experimental research methods were used. HSFs were collected and divided into 200 mV/mm electric field group treated with 200 mV/mm electric field for 6 h and simulated electric field group placed in the electric field device without electricity for 6 h. Changes in morphology and arrangement of cells were observed in the living cell workstation; the number of cells at 0 and 6 h of treatment was recorded, and the rate of change in cell number was calculated; the direction of cell movement, movement velocity, and trajectory velocity within 3 h were observed and calculated (the number of samples was 34 in the simulated electric field group and 30 in 200 mV/mm electric field group in the aforementioned experiments); the protein expression of α-smooth muscle actin (α-SMA) in cells after 3 h of treatment was detected by immunofluorescence method (the number of sample was 3). HSFs were collected and divided into simulated electric field group placed in the electric field device without electricity for 3 h, and 100 mV/mm electric field group, 200 mV/mm electric field group, and 400 mV/mm electric field group which were treated with electric fields of corresponding intensities for 3 h. Besides, HSFs were divided into simulated electric field group placed in the electric field device without electricity for 6 h, and electric field treatment 1 h group, electric field treatment 3 h group, and electric field treatment 6 h group treated with 200 mV/mm electric field for corresponding time. The protein expressions of α-SMA and proliferating cell nuclear antigen (PCNA) were detected by Western blotting (the number of sample was 3). Data were statistically analyzed with Mann-Whitney U test, one-way analysis of variance, independent sample t test, and least significant difference test. Results After 6 h of treatment, compared with that in simulated electric field group, the cells in 200 mV/mm electric field group were elongated in shape and locally adhered; the cells in simulated electric field group were randomly arranged, while the cells in 200 mV/mm electric field group were arranged in a regular longitudinal direction; the change rates in the number of cells in the two groups were similar (P>0.05). Within 3 h of treatment, the cells in 200 mV/mm electric field group had an obvious tendency to move toward the positive electrode, and the cells in simulated electric field group moved around the origin; compared with those in simulated electric field group, the movement velocity and trajectory velocity of the cells in 200 mV/mm electric field group were increased significantly (with Z values of -5.33 and -5.41, respectively, P<0.01), and the directionality was significantly enhanced (Z=-4.39, P<0.01). After 3 h of treatment, the protein expression of α-SMA of cells in 200 mV/mm electric field group was significantly higher than that in simulated electric field group (t=-9.81, P<0.01). After 3 h of treatment, the protein expressions of α-SMA of cells in 100 mV/mm electric field group, 200 mV/mm electric field group, and 400 mV/mm electric field group were 1.195±0.057, 1.606±0.041, and 1.616±0.039, respectively, which were significantly more than 0.649±0.028 in simulated electric field group (P<0.01). Compared with that in 100 mV/mm electric field group, the protein expressions of α-SMA of cells in 200 mV/mm electric field group and 400 mV/mm electric field group were significantly increased (P<0.01). The protein expressions of α-SMA of cells in electric field treatment 1 h group, electric field treatment 3 h group, and electric field treatment 6 h group were 0.730±0.032, 1.561±0.031, and 1.553±0.045, respectively, significantly more than 0.464±0.020 in simulated electric field group (P<0.01). Compared with that in electric field treatment 1 h group, the protein expressions of α-SMA in electric field treatment 3 h group and electric field treatment 6 h group were significantly increased (P<0.01). After 3 h of treatment, compared with that in simulated electric field group, the protein expressions of PCNA of cells in 100 mV/mm electric field group, 200 mV/mm electric field group, and 400 mV/mm electric field group were significantly decreased (P<0.05 or P<0.01); compared with that in 100 mV/mm electric field group, the protein expressions of PCNA of cells in 200 mV/mm electric field group and 400 mV/mm electric field group were significantly decreased (P<0.05 or P<0.01); compared with that in 200 mV/mm electric field group, the protein expression of PCNA of cells in 400 mV/mm electric field group was significantly decreased (P<0.01). Compared with that in simulated electric field group, the protein expressions of PCNA of cells in electric field treatment 1 h group, electric field treatment 3 h group, and electric field treatment 6 h group were significantly decreased (P<0.01); compared with that in electric field treatment 1 h group, the protein expressions of PCNA of cells in electric field treatment 3 h group and electric field treatment 6 h group were significantly decreased (P<0.05 or P<0.01); compared with that in electric field treatment 3 h group, the protein expression of PCNA of cells in electric field treatment 6 h group was significantly decreased (P<0.01). Conclusions The bio-intensity electric field can induce the migration of HSFs and promote the transformation of fibroblasts to myofibroblasts, and the transformation displays certain dependence on the time and intensity of electric field. -
Key words:
- Skin /
- Fibroblasts /
- Myofibroblasts /
- Cell proliferation /
- Cell transformation /
- Bio-intensity electric fields
-
(1)通过改进胰管穿刺置管法获取胆胰混合液。
(2)证实严重烫伤大鼠伤后早期胰腺外分泌功能下降。
(3)观察到大鼠烫伤面积越大,胰腺外分泌功能损伤越严重,并可能出现胰腺炎的早期病理改变。
肠内营养的有效吸收依赖于多种消化酶的正常分泌和功能行使。胰酶是一种重要的消化酶,主要含胰脂肪酶、α-淀粉酶和胰蛋白酶等。一方面,胰酶分泌的生理过程反映了胰腺外分泌功能(PEF)的状态;另一方面,以酶原形式存在的胰酶被不当或过早激活后会表现为胰腺炎,这2种表现在严重烧伤后发生与否及其变化规律目前鲜有报道。由各种原因引起的胰酶分泌不足或胰酶分泌不同步,或肠腔胰酶活性降低导致患者出现的营养消化吸收不良等症状被称为胰腺外分泌功能不全(PEI) [ 1] 。2016年,国际烧伤学会在烧伤救治实践指南中强调营养评估和管理的重要性,并提出应尽早(伤后24 h内,甚至在伤后6~12 h内)对烧伤患者实施肠内营养 [ 2, 3] ,以恢复和保护肠道功能,防止过多毒素入血,保护胃肠黏膜,减少胃肠道并发症 [ 4] 。目前,PEF的常规检测在烧伤患者中并未开展。胆囊收缩素(cholecystokinin,CCK)/促胰液素/蛙皮素是常见的胰岛素分泌促进剂,通常采用生理剂量的分泌促进剂刺激胰酶,检测PEF [ 5] 。本研究拟通过构建2种面积严重烫伤大鼠模型,在PEF的常规检测方法上加以改进,用以研究严重烫伤大鼠早期合成和分泌胰酶能力的变化规律以及胰腺外分泌的组织损伤和修复规律,以期为临床防治严重烧伤后胰腺相关疾病和制订营养支持方案提供理论支持。
1. 材料与方法
本实验研究遵循解放军总医院实验动物伦理审查委员会和国家有关实验动物管理使用的规定。
1.1 动物及主要试剂与仪器来源
80只清洁级7~8周龄雄性健康SD大鼠,体重(230±20)g,购自北京科宇动物养殖中心,许可证号:SCXK(京)2018-0010。
CCK八肽(CCK8)购自大连美仑生物技术有限公司,大鼠α-淀粉酶和胰脂肪酶活力检测试剂盒购自南京建成生物工程研究所,大鼠α-淀粉酶、胰脂肪酶、胰蛋白酶、IL-1β和IL-6的ELISA试剂盒购自上海酶联生物科技有限公司,兔抗大鼠缺氧诱导因子1α(HIF-1α)抗体购自北京博奥森生物技术有限公司,Alexa Fluor 488标记的山羊抗兔IgG抗体购自北京百奥莱博科技有限公司。
RT-6100型酶标仪购自美国Rayto公司,CKX53型倒置荧光显微镜购自日本Olympus公司,H-7650型透射电子显微镜购自日本日立公司。
1.2 动物模型制备及分组与处理
取80只大鼠,按随机数字表法分为单纯假伤组、假伤+CCK8组、严重烫伤+CCK8组、特重烫伤+CCK8组,前2组各8只、后2组各32只。各组大鼠于伤前禁食12 h,并于烫伤前2 h按照1 mL/kg的剂量经口腔注入液体石蜡以排净粪便。将2组假伤大鼠置于37 ℃温水中作假伤处理,将假伤+CCK8组大鼠伤后即刻右侧股静脉插入留置针并用丝线固定,按照80 pmol·L -1·kg -1·h -1剂量泵入0.1 g/L的CCK8溶液(用含体积分数1%胎牛血清的生理盐水配制)进行刺激,每只大鼠仅刺激1次。后2组大鼠按照文献[ 6]的方法分别在制作30%TBSA和50%TBSA的严重/特重烫伤模型后进行常规抗休克处理,创面每日碘伏消毒;伤后1、2、3、7 d,每个时间点选取8只大鼠同前给予CCK8刺激,每只大鼠仅刺激1次,且刺激前均禁食12 h。
1.3 PEF检测
1.3.1 胆胰混合液(PBJ)的收集与胰酶含量检测
采用改进的胰管穿刺置管法动态收集PBJ(淡黄色)。大鼠常规麻醉后,于剑突下1 cm处腹部正中逐层剪开皮肤、腹肌并分离至腹腔,确定散在分布的大鼠胰腺组织,顺十二指肠降部找到十二指肠乳头及透明胰管,于贴近十二指肠乳头处从胰管外壁插入F22密闭式防刺伤静脉留置针,留置针后接微量收集器,见 图1。单纯假伤组大鼠的PBJ收集在伤后即刻进行,假伤+CCK8组、严重烫伤+CCK8组、特重烫伤+CCK8组大鼠采用边同1.2 CCK8刺激边收集PBJ的方式,持续收集1 h并记录液体体积,收集过程中整体收集装置置于冰上。PBJ的分泌体积可反映胰管分泌胰酶和水的能力。样本数为8。
按照胰脂肪酶、α-淀粉酶和胰蛋白酶的ELISA试剂盒的说明书,采用酶标仪测定波长450 nm处的吸光度值,反映1 h内PBJ分泌体积中胰脂肪酶、α-淀粉酶和胰蛋白酶的浓度并计算其含量,胰酶各组分含量=胰酶各组分浓度×PBJ分泌体积。样本数为8。
1.3.2 血液收集及其所含胰酶活力检测
PBJ收集完成后即刻,于大鼠左侧股静脉收集血液约2 mL,室温下静置2 h,待分层后于4 ℃离心机中以离心半径8 cm、3 000 r/min离心10 min。吸取上清液,采用标准比色法检测血清中胰脂肪酶和α-淀粉酶的浓度,反映其活力。为保持胰腺组织形态,避免置管导致胰腺炎 [ 7] ,微量收集器全程保持负压值为-0.3 kPa。样本数为8。
1.4 胰腺组织损伤检测
完成1.3中2种液体收集后即刻,沿胰管走行、胃及十二指肠内侧和脾门之间剪取散在分布的胰腺组织,一部分保存于-80 ℃冰箱、一部分用石蜡包埋、一部分浸泡于40 g/L的多聚甲醛溶液中固定、一部分新鲜组织浸泡于PBS中备用。
1.4.1 组织炎症因子水平检测
取冰冻的胰腺组织,按照IL-1β和IL-6的ELISA试剂盒说明书,采用酶标仪测定波长450 nm处的吸光度值,反映其浓度。样本数为8。
1.4.2 胰腺缺氧损伤水平检测
取石蜡包埋的胰腺组织,采用免疫荧光法检测组织中HIF-1α的表达,一抗为兔抗大鼠HIF-1α抗体(稀释比为1∶400)、二抗为Alexa Fluor 488标记的山羊抗兔IgG抗体(稀释比为1∶1 000)。采用4',6-二脒基-2-苯基吲哚染核后,于100倍倒置荧光显微镜下采集图像。样本数为3。
1.4.3 组织病理学检测
取多聚甲醛溶液中固定的胰腺组织,进行常规HE染色。同时,采用改良Schmidt法 [ 8] 评估2组烫伤大鼠胰腺组织损伤的严重程度,包含水肿、炎症浸润、出血、变性坏死情况的4个参数。在40倍视野下任意选取6张图片评估各参数得分,反映胰腺损伤的严重程度,每个变量的评估如下。(1)水肿:无水肿为0分,小叶间间隙局部水肿为1分,小叶内间隙均出现水肿为2分,胰腺腺泡呈孤立岛状水肿为3分;(2)出血:无出血为0分,轻度出血为1分,中度出血为2分,重度出血为3分;(3)变性坏死:腺泡细胞局灶性变性坏死(面积<5%)为1分,和/或小叶下腺泡细胞变性坏死(面积<20%)为2分,和/或小叶腺泡细胞变性坏死(面积≥20%)为3分;(4)炎症浸润:无炎症为0分,轻度炎症为1分,中度炎症为2分,重度炎症为3分。得分越高,反映组织损伤程度越严重。样本数为6。
1.4.4 腺泡细胞超微结构
参照文献[ 9],取PBS中新鲜胰腺组织,经固定、脱水、渗透包埋、聚合、切片、染色后,使用透射电子显微镜观察并采集5 000倍镜下图像,观察以下内容:(1)腺泡细胞形态变化以及细胞间连接。(2)腺泡细胞内酶原颗粒大小、数量及形态变化,特别是管腔侧(顶端膜)及基底侧(基底膜)酶原颗粒数量及形态特征变化。
1.5 统计学处理
采用SPSS 26.0统计软件和Graphpad Prism 9.0统计软件进行数据分析。计量资料数据均符合正态分布,以
2. 结果
2.1 胰腺组织分泌PBJ体积
与单纯假伤组相比,假伤+CCK8组大鼠伤后即刻胰腺组织1 h内的PBJ分泌体积显著升高( t=4.92, P<0.001)。与假伤+CCK8组伤后即刻相比,严重烫伤+CCK8组、特重烫伤+CCK8组大鼠伤后1、2、3、7 d胰腺组织1 h内的PBJ分泌体积均显著下降( P<0.001)。与严重烫伤+CCK8组相比,特重烫伤+CCK8组大鼠伤后1、2 d胰腺组织1 h内的PBJ分泌体积均显著下降( P<0.001)。其中,严重烫伤+CCK8组大鼠胰腺组织1 h内的PBJ分泌体积于伤后3 d降至最低,特重烫伤+CCK8组大鼠胰腺组织1 h内的PBJ分泌体积于伤后2 d降至最低。见 表1。
表1 2组假伤大鼠伤后即刻和2组烫伤大鼠伤后不同时间点胰腺组织外分泌功能及炎症因子水平的比较(组别与时间点 样本数 1 h内PBJ分泌体积(mL) 胰酶含量 血清中胰酶活力(U/L) 胰腺组织中炎症因子水平(pg/g) 胰脂肪酶(ng) α-淀粉酶(ng) 胰蛋白酶(pg) 胰脂肪酶 α-淀粉酶 IL-1β IL-6 单纯假伤组 8 伤后即刻 0.740±0.030 65.1±3.9 20.7±1.8 932±96 0.81±0.04 637±44 269±15 658±17 假伤+CCK8组 8 伤后即刻 0.823±0.033 a 73.3±2.7 a 23.2±1.8 a 1 184±93 a 0.86±0.08 657±25 265±9 649±31 严重烫伤+CCK8组 32 伤后1 d 0.681±0.024 b 61.0±2.2 b 20.3±1.1 b 1 262±52 0.88±0.22 641±16 291±7 b 1 060±95 b 伤后2 d 0.608±0.056 b 60.0±1.7 b 20.2±1.0 b 1 226±113 0.96±0.13 647±17 349±10 b 1 236±12 b 伤后3 d 0.525±0.025 b 52.0±2.5 b 17.0±0.8 b 1 059±83 1.00±0.22 651±17 381±15 b 1 316±63 b 伤后7 d 0.720±0.044 b 64.1±2.0 b 20.2±0.8 b 1 131±139 0.88±0.14 662±22 285±14 994±95 b 特重烫伤+CCK8组 32 伤后1 d 0.540±0.025 bc 51.6±2.7 bc 17.7±0.7 bc 1 094±93 c 1.13±0.31 654±31 303±12 b 1 178±136 b 伤后2 d 0.406±0.021 bc 41.7±1.3 bc 14.4±0.6 bc 947±67 bc 1.16±0.25 666±32 408±14 bc 1 487±183 bc 伤后3 d 0.475±0.036 b 43.8±2.0 bc 15.2±0.9 b 1 056±116 1.13±0.31 670±17 403±12 bc 1 390±28 b 伤后7 d 0.690±0.018 b 66.2±2.6 b 20.9±1.2 b 1 190±73 0.98±0.22 679±8 294±13 b 1 010±103 b 注:假伤指于37 ℃温水中模拟烫伤,严重烫伤指30%体表总面积(TBSA)烫伤,特重烫伤指50%TBSA烫伤;CCK8为胆囊收缩素八肽,PBJ为胆胰混合液,IL为白细胞介素;2个烫伤组各时间点样本数均为8;胰酶含量为1 h内PBJ分泌体积中含量;1 h内PBJ分泌体积,胰脂肪酶、α-淀粉酶、胰蛋白酶含量,血清中胰脂肪酶、α-淀粉酶活力,胰腺组织中IL-1β、IL-6水平的处理因素主效应, F值分别为566.18与700.91、209.42、11.60及10.99、2.88和442.72、329.04, P值分别为<0.001与<0.001、<0.001、<0.001及<0.001、0.127和<0.001、<0.001;时间因素主效应, F值分别为79.89与104.35、21.80、3.46及0.87、1.95和208.62、34.90, P值分别为<0.001与<0.001、<0.001、0.020及0.461、0.127和<0.001、<0.001;两者交互作用, F值分别为30.15与49.75、12.99、5.49及0.45、0.55和59.51、10.90, P值分别为<0.001与<0.001、<0.001、<0.001及0.846、0.766和<0.001、<0.001;与单纯假伤组相比, a P<0.05;与假伤+CCK8组相比, b P<0.05;与严重烫伤+CCK8组相比, c P<0.05 2.2 PBJ中的胰酶含量
相较于单纯假伤组伤后即刻,假伤+CCK8组大鼠伤后即刻PBJ中胰脂肪酶、α-淀粉酶和胰蛋白酶含量均显著升高( t值分别为4.56、3.30、4.99, P值分别为<0.001、0.005、<0.001)。与假伤+CCK8组伤后即刻相比,严重烫伤+CCK8组和特重烫伤+CCK8组大鼠伤后1、2、3、7 d的PBJ中胰脂肪酶和α-淀粉酶含量均显著降低( P<0.001),特重烫伤+CCK8组大鼠伤后2 d的PBJ中胰蛋白酶含量显著降低( P<0.001)。与严重烫伤+CCK8组相比,特重烫伤+CCK8组大鼠伤后1、2、3 d的PBJ中胰脂肪酶含量均显著降低( P<0.001),伤后1、2 d的PBJ中α-淀粉酶含量( P值分别为0.003、<0.001)和胰蛋白酶含量( P值分别为0.049、<0.001)均显著降低。见 表1。
2.3 血清中的胰酶活力
与单纯假伤组伤后即刻相比,假伤+CCK8组伤后即刻大鼠血清中的胰脂肪酶活力无明显变化( t=1.24, P=0.234)。与假伤+CCK8组伤后即刻相比,严重烫伤+CCK8组和特重烫伤+CCK8组大鼠伤后1、2、3、7 d血清中的胰脂肪酶活力均无明显变化( P>0.05)。4组大鼠伤后各时间点血清中α-淀粉酶活力比较,差异均无统计学意义( P>0.05)。见 表1。
2.4 组织炎症因子水平
相较于单纯假伤组伤后即刻,假伤+CCK8组大鼠伤后即刻血清中的IL-1β和IL-6水平均无明显变化( t值分别为0.69和0.65, P值分别为0.504和0.528)。与假伤+CCK8组大鼠伤后即刻相比,严重烫伤+CCK8组大鼠伤后1、2、3 d和特重烫伤+CCK8组大鼠伤后1、2、3、7 d胰腺组织中的IL-1β水平均显著升高( P值分别为0.006、<0.001、<0.001,<0.001、<0.001、<0.001、0.001),严重烫伤+CCK8组和特重烫伤+CCK8组大鼠伤后1、2、3、7 d胰腺组织中的IL-6水平均显著升高( P<0.001)。与严重烫伤+CCK8组相比,特重烫伤+CCK8组大鼠伤后2、3 d胰腺组织中的IL-1β水平均显著升高( P值分别为<0.001、0.031),伤后2 d胰腺组织中的IL-6水平显著升高( P<0.001)。见 表1。
2.5 组织缺氧损伤水平
单纯假伤组和假伤+CCK8组大鼠伤后即刻胰腺组织中HIF-1α表达水平均较低。相较于假伤+CCK8组大鼠伤后即刻,2组烫伤大鼠胰腺组织中HIF-1α表达水平于伤后不同时间点均呈一定程度升高,其表达位置由胰腺组织边缘向全胰腺过渡。伤后7 d,2组烫伤大鼠胰腺组织中HIF-1α表达水平下降且趋于正常。见 图2。
 注:假伤指于37 ℃温水中模拟烫伤,严重烫伤指30%体表总面积(TBSA)烫伤,特重烫伤指50%TBSA烫伤;CCK8为胆囊收缩素八肽;缺氧诱导因子1α(HIF-1α)阳性染色为绿色(反映组织缺氧损伤程度),细胞核阳性染色为蓝色
注:假伤指于37 ℃温水中模拟烫伤,严重烫伤指30%体表总面积(TBSA)烫伤,特重烫伤指50%TBSA烫伤;CCK8为胆囊收缩素八肽;缺氧诱导因子1α(HIF-1α)阳性染色为绿色(反映组织缺氧损伤程度),细胞核阳性染色为蓝色2.6 组织病理学情况
单纯假伤组大鼠伤后即刻胰腺组织结构正常,小叶间隙清晰,视野中腺泡结构正常,未见坏死,小叶内导管上皮细胞排列整齐,未见异常,胰岛结构清晰。假伤+CCK8组大鼠伤后即刻胰腺组织结构与单纯假伤组近似,但腺泡细胞胞质占比增大。严重烫伤+CCK8组大鼠伤后1 d,胰腺组织结构异常,间质水肿,视野中可见多处腺泡细胞坏死,部分小叶内导管细胞排列不齐,管腔结构消失;伤后2 d,胰腺组织结构重度异常,腺泡细胞呈片状坏死,且主要存在于胰腺边缘部位,视野中可见组织大面积出血和水肿,个别小叶内导管上皮细胞点状坏死,部分组织间隙可见中性粒细胞浸润;伤后3 d,组织结构重度异常,组织结构紊乱,仅存少量结构清晰的腺泡,小叶内导管细胞结构紊乱,管腔结构基本消失,细胞形态和组织关系已基本无法辨认,视野中可见大面积腺泡细胞坏死及少量炎症细胞浸润;伤后7 d,组织结构基本恢复正常,未见出血及炎症细胞浸润,腺泡葡萄状结构基本清晰,未见坏死,小叶内导管细胞排列整齐,管腔结构基本恢复。其中,腺泡细胞的坏死灶主要出现在胰腺周边部分,而导管细胞的结构紊乱及坏死散在分布于整个胰腺小叶。特重烫伤+CCK8组相较于严重烫伤+CCK8组大鼠表现出更早期和更严重的胰腺组织损伤。见 图3。
 注:假伤指于37 ℃温水中模拟烫伤,严重烫伤指30%体表总面积(TBSA)烫伤,特重烫伤指50%TBSA烫伤;CCK8为胆囊收缩素八肽;黄色箭头指示小叶间导管细胞,蓝色箭头指示炎症细胞
注:假伤指于37 ℃温水中模拟烫伤,严重烫伤指30%体表总面积(TBSA)烫伤,特重烫伤指50%TBSA烫伤;CCK8为胆囊收缩素八肽;黄色箭头指示小叶间导管细胞,蓝色箭头指示炎症细胞与严重烫伤+CCK8组相比,特重烫伤+CCK8组大鼠伤后各时间点胰腺组织水肿、炎症细胞浸润、出血和坏死得分表现出不同程度的升高。此外,2组烫伤大鼠胰腺组织各项目评分均于伤后7 d基本恢复正常。见 图4。
2.7 胰泡细胞超微结构
单纯假伤组大鼠伤后即刻胰腺组织结构正常,腺泡细胞间隙清晰,胞内酶原颗粒染色淡且数量较少,细胞核结构正常。假伤+CCK8组大鼠伤后即刻胰腺组织可见腺泡细胞内密集分布的致密酶原颗粒,顶端分泌增强。严重烫伤+CCK8组大鼠伤后1、2、3 d胰腺组织腺泡细胞间隙逐渐变窄,结构不清,细胞内酶原颗粒数量较假伤+CCK8组伤后即刻少,细胞核形态不规则、分裂边集;伤后7 d,胰腺组织腺泡细胞间隙清晰,细胞核及小叶内导管管腔结构正常,腺泡细胞极性分泌逐渐恢复,酶原颗粒数量及密度亦逐渐恢复。特重烫伤+CCK8组相较于严重烫伤+CCK8组大鼠表现出更早期和更严重的腺泡细胞损伤。见 图5。
 注:假伤指于37 ℃温水中模拟烫伤,严重烫伤指30%体表总面积(TBSA)烫伤,特重烫伤指50%TBSA烫伤;CCK8为胆囊收缩素八肽;绿色箭头指示酶原颗粒,蓝色箭头指示细胞核
注:假伤指于37 ℃温水中模拟烫伤,严重烫伤指30%体表总面积(TBSA)烫伤,特重烫伤指50%TBSA烫伤;CCK8为胆囊收缩素八肽;绿色箭头指示酶原颗粒,蓝色箭头指示细胞核3. 讨论
胰腺原发创伤、外科切除,以及急、慢性胰腺炎 [ 10, 11] 是PEF下降最主要的原因。采用促泌素刺激胰腺组织获取胰液进行酶学分析是评估PEF最常用的方法,其在轻度PEI的诊断中应用价值较高。为避免胰管内穿刺置管的创伤、复杂操作及内镜置管带来的其他风险,如经内镜逆行胰胆管造影操作可能引起小肠液逆流入胰管内导致胰酶过早激活 [ 12] 并引起胰管内压升高 [ 7] ,本研究将置管入口改为胰管外侧壁,避免损伤奥迪括约肌,且胰液收集装置保持一定负压,可进一步减少因穿刺置管增加的胰腺炎风险。
本实验中,严重烫伤大鼠胰酶合成和分泌均受到显著抑制,这一变化可能与缺血缺氧损伤和炎症打击有关。严重烧伤早期机体心功能下降和相对血容量低,增加了组织的损伤程度和器官的功能障碍 [ 13] ,这一过程一般持续1~2 d。严重烧伤后1~3 d进入高代谢和高动力阶段 [ 14] 。代谢紊乱、细菌移位等因素也可能引起或加重胰腺损伤 [ 15] 。然而,目前对于严重烧伤后胰腺腺泡细胞和导管细胞的研究匮乏。本实验结果显示,严重烫伤大鼠急性期PEF不同程度受损,且伤后2~3 d受损程度最重,表现为腺泡细胞合成胰酶减少,导管细胞分泌水的严重减少,胰腺组织不同程度水肿、出血及坏死。此外,严重烫伤后腺泡细胞的损伤源于胰腺外周,而导管细胞的坏死散在分布于整个胰腺小叶。本研究还显示,与缺氧损伤直接相关的HIF-1α蛋白表达于伤后急性期内显著升高,且烫伤面积越大,其表达水平越高。此外,烫伤大鼠体内释放大量炎症介质可进一步加重胰腺血管内皮损伤 [ 13] ,这可能是严重烫伤后大鼠PEF下降的另一个重要原因。严重烫伤大鼠胰腺组织中IL-1β和IL-6水平于急性期达到峰值;随着炎症因子水平下降,PEF的损伤程度也得到恢复。PEF的变化规律与严重烧伤引起的缺血缺氧打击基本一致,但其损伤机制还有待进一步研究。
PEF损伤的形式除胰酶合成和分泌减少外,还表现为胰酶分泌方式的紊乱,如胞内囊泡融合、极化运输、胞吐和导管内胰液运输等过程的异常变化,该变化提示可能发生了胰腺炎 [ 16] 。而由胰酶分泌方式紊乱导致的胰腺炎的诊断治疗过程与PEI完全不同 [ 1, 17] 。本研究中,严重烫伤除导致胰酶合成和分泌减少外,还可见部分腺泡细胞内酶原颗粒极性运输发生紊乱,并出现自噬泡,提示部分腺泡细胞内出现了胰腺炎的早期事件 [ 9, 18] ,且以上现象在特重烫伤大鼠胰腺组织中表现更明显。患者烧伤后继发胰腺炎的现象常被当作个案报道,但其在儿童和老年烧伤患者中发生率较高 [ 19, 20] ,猜测可能与肠上皮黏膜屏障功能下降引起细菌移位有关。因此,要防治特大面积烧伤患者继发胰腺炎。
胰腺外分泌部在一定条件下可以实现再生,例如创伤或胰腺切除术后的腺泡细胞再生主要来源于剩余胰腺组织 [ 21] ,胰腺损伤情况下,位于腺泡周围和导管周围的胰腺星状细胞可被激活分化为腺泡细胞 [ 22] 。Criscimanna等 [ 23] 使用白喉毒素诱导小鼠腺泡细胞死亡时观察到,腺泡细胞的再生是由导管分化实现的,而非腺泡自我更新。本实验显示,烫伤大鼠胰腺导管细胞的损伤程度显著重于腺泡细胞,可能会影响到导管到腺泡的转化。胰腺内外分泌部之间的联系不可分割,1型糖尿病或极端条件下导致缺失的胰岛β细胞不仅可由胰岛α、δ细胞分化而来,也可由胰腺外分泌部的导管细胞和腺泡细胞转化而来 [ 24, 25] 。高糖状态下,胰腺外分泌细胞的再生受到抑制,有效控制了血糖水平,维持了内环境的稳定,从而减轻创伤导致的胰腺外分泌损伤 [ 26] 。
对患者实施早期肠内营养具有诸多益处,如可促进胃泌素、CCK等的分泌,还可促进胃肠蠕动、降低肠道通透性和维持肠黏膜屏障功能,以及降低感染相关的并发症。本研究中,严重烫伤大鼠急性期内胰酶合成和分泌显著下降,提示在临床中要重视肠内营养给予途径和营养素配比。此外,烧伤患者发生肠内喂养不耐受的风险较高,而不当的肠内营养方式或许会导致较差的临床结局。目前的观点认为,应对中重度胰腺炎患者实施肠内喂养,且空肠喂养仍然是肠内营养的首选途径 [ 27] 。小肠扩张可导致胰腺炎,可能与压力升高引起激素变化 [ 28] 及胰管内压升高 [ 7] 有关。因此,严重烧伤患者急性期内的肠内营养途径宜选择鼻空肠管和肠外营养相结合的方式,以应对PEF下降,避免发生腹泻及胰腺炎。
烧伤机体的高代谢反应使得患者在急性期内乃至长期对能量和营养的需求剧增,而蛋白质消化产物(如肽和氨基酸)和脂肪消化产物(如脂肪酸和单甘酯)又可有效促进胰腺的外分泌。本研究显示,烫伤大鼠较假伤大鼠的胰脂肪酶和α-淀粉酶含量均基本呈不同程度下降趋势,而胰蛋白酶仅于急性期下降,提示临床上在严重烧伤患者急性期内给予肠内营养时可减少长链脂肪酸、多肽类占比,并增加无须消化的氨基酸和短肽类。此外,CCK不仅可以促进胰酶的合成及分泌,还通过与CCK受体结合以及刺激迷走神经实现抗炎作用,因此对由炎症诱导的肠通透性变化可通过切断迷走神经和给予CCK和烟碱受体拮抗剂消除 [ 29] ,烧伤急性期给予CCK治疗或许可以帮助患者改善PEF。然而,大剂量的CCK可导致急性胰腺炎 [ 30] ,可能机制为CCK受体的不同亚基构象发生变化会激活不同的下游信号,从而引起细胞质钙离子水平超负荷。因此,对烧伤患者行CCK干预治疗的有效性和安全性还须进一步研究。
本研究揭示了烧伤后早期PEF的变化规律,证实了严重烧伤可影响胰酶的合成和分泌,但其机制还须进一步研究。
王文平:设计实验、实施实验、采集数据、分析数据、撰写文章;冀然、邬亚婷:实施实验、采集数据;张泽:实施实验、采集数据、分析数据;张恒术:撰写并审阅文章,获取研究经费,行政、技术和材料支持;张琼:分析数据、技术和材料支持;江旭品、滕苗:设计实验,撰写并审阅文章,获取研究经费,行政、技术和材料支持所有作者均声明不存在利益冲突 -
参考文献
(40) [1] JiR,TengM,ZhangZ,et al.Electric field down-regulates CD9 to promote keratinocytes migration through AMPK pathway[J].Int J Med Sci,2020,17(7):865-873.DOI: 10.7150/ijms.42840. [2] YanT,JiangX,GuoX,et al.Electric field-induced suppression of PTEN drives epithelial-to-mesenchymal transition via mTORC1 activation[J].J Dermatol Sci,2017,85(2):96-105.DOI: 10.1016/j.jdermsci.2016.11.007. [3] BostanLE,AlmqvistS,PullarCE.A pulsed current electric field alters protein expression creating a wound healing phenotype in human skin cells[J].Regen Med,2020,15(5):1611-1623.DOI: 10.2217/rme-2019-0087. [4] NuccitelliR.A role for endogenous electric fields in wound healing[J].Curr Top Dev Biol,2003,58:1-26.DOI: 10.1016/s0070-2153(03)58001-2. [5] AbeR,DonnellySC,PengT,et al.Peripheral blood fibrocytes: differentiation pathway and migration to wound sites[J].J Immunol,2001,166(12):7556-7562.DOI: 10.4049/jimmunol.166.12.7556. [6] DeesC,ChakrabortyD,DistlerJHW.Cellular and molecular mechanisms in fibrosis[J].Exp Dermatol,2021,30(1):121-131.DOI: 10.1111/exd.14193. [7] RemstDF,Blaney DavidsonEN,van der KraanPM.Unravelling osteoarthritis-related synovial fibrosis: a step closer to solving joint stiffness[J].Rheumatology (Oxford),2015,54(11):1954-1963.DOI: 10.1093/rheumatology/kev228. [8] KamilS,MohanRR.Corneal stromal wound healing: major regulators and therapeutic targets[J].Ocul Surf,2021,19:290-306.DOI: 10.1016/j.jtos.2020.10.006. [9] 刘杰,任淅,郭小伟,等.直流电场对BALB/c小鼠乳鼠真皮成纤维细胞定向迁移与排列的作用及其机制[J].中华烧伤杂志,2016,32(4):224-231.DOI: 10.3760/cma.j.issn.1009-2587.2016.04.007. [10] 冀然,张泽,王文平,等.生物强度电场对人表皮细胞株HaCaT和小鼠表皮细胞运动性及CD9表达的影响[J].中华烧伤杂志,2021,37(1):34-41.DOI: 10.3760/cma.j.cn501120-20200115-00023. [11] TredgetEE,LeviB,DonelanMB.Biology and principles of scar management and burn reconstruction[J].Surg Clin North Am,2014,94(4):793-815.DOI: 10.1016/j.suc.2014.05.005. [12] BarrettLW,FearVS,WaithmanJC,et al.Understanding acute burn injury as a chronic disease[J/OL].Burns Trauma,2019,7:23[2022-03-16].https://pubmed.ncbi.nlm.nih.gov/31534977/. DOI: 10.1186/s41038-019-0163-2. [13] DesmoulièreA,DarbyIA,GabbianiG.Normal and pathologic soft tissue remodeling: role of the myofibroblast, with special emphasis on liver and kidney fibrosis[J].Lab Invest,2003,83(12):1689-1707.DOI: 10.1097/01.lab.0000101911.53973.90. [14] CulleyOJ,LouisB,PhilippeosC,et al.Differential expression of insulin-like growth factor 1 and Wnt family member 4 correlates with functional heterogeneity of human dermal fibroblasts[J].Front Cell Dev Biol,2021,9:628039.DOI: 10.3389/fcell.2021.628039. [15] TomasekJJ,GabbianiG,HinzB,et al.Myofibroblasts and mechano-regulation of connective tissue remodelling[J].Nat Rev Mol Cell Biol,2002,3(5):349-363.DOI: 10.1038/nrm809. [16] XinY,MinP,XuH,et al.CD26 upregulates proliferation and invasion in keloid fibroblasts through an IGF-1-induced PI3K/AKT/mTOR pathway[J/OL].Burns Trauma,2020,8:tkaa025[2022-03-16].https://pubmed.ncbi.nlm.nih.gov/33150188/. DOI: 10.1093/burnst/tkaa025. [17] ModarressiA,PietramaggioriG,GodboutC,et al.Hypoxia impairs skin myofibroblast differentiation and function[J].J Invest Dermatol,2010,130(12):2818-2827.DOI: 10.1038/jid.2010.224. [18] Demidova-RiceTN,HamblinMR,HermanIM.Acute and impaired wound healing: pathophysiology and current methods for drug delivery, part 2: role of growth factors in normal and pathological wound healing: therapeutic potential and methods of delivery[J].Adv Skin Wound Care,2012,25(8):349-370.DOI: 10.1097/01.ASW.0000418541.31366.a3. [19] PutnikP,KresojaŽ,BosiljkovT,et al.Comparing the effects of thermal and non-thermal technologies on pomegranate juice quality: a review[J].Food Chem,2019,279:150-161.DOI: 10.1016/j.foodchem.2018.11.131. [20] SnyderS,DeJuliusC,WillitsRK.Electrical stimulation increases random migration of human dermal fibroblasts[J].Ann Biomed Eng,2017,45(9):2049-2060.DOI: 10.1007/s10439-017-1849-x. [21] SuessPM,SmithSA,MorrisseyJH.Platelet polyphosphate induces fibroblast chemotaxis and myofibroblast differentiation[J].J Thromb Haemost,2020,18(11):3043-3052.DOI: 10.1111/jth.15066. [22] NguyenEB,WishnerJ,SlowinskaK.The effect of pulsed electric field on expression of ECM proteins: collagen, elastin, and MMP1 in human dermal fibroblasts[J].J Electroanal Chem (Lausanne),2018,812:265-272.DOI: 10.1016/j.jelechem.2018.01.050. [23] ChaponnierC,GabbianiG.Pathological situations characterized by altered actin isoform expression[J].J Pathol,2004,204(4):386-395.DOI: 10.1002/path.1635. [24] FroidureA,Marchal-DuvalE,Homps-LegrandM,et al.Chaotic activation of developmental signalling pathways drives idiopathic pulmonary fibrosis[J].Eur Respir Rev,2020,29(158):190140. DOI: 10.1183/16000617.0140-2019. [25] Vaamonde-GarciaC,MalaiseO,CharlierE,et al.15-Deoxy-Δ-12, 14-prostaglandin J2 acts cooperatively with prednisolone to reduce TGF-β-induced pro-fibrotic pathways in human osteoarthritis fibroblasts[J].Biochem Pharmacol,2019,165:66-78.DOI: 10.1016/j.bcp.2019.03.039. [26] SidgwickGP,BayatA.Extracellular matrix molecules implicated in hypertrophic and keloid scarring[J].J Eur Acad Dermatol Venereol,2012,26(2):141-152.DOI: 10.1111/j.1468-3083.2011.04200.x. [27] TanJ,WuJ.Current progress in understanding the molecular pathogenesis of burn scar contracture[J/OL].Burns Trauma,2017,5:14[2022-03-16]. https://pubmed.ncbi.nlm.nih.gov/28546987/. DOI: 10.1186/s41038-017-0080-1. [28] WangL,KongW,LiuB,et al.Proliferating cell nuclear antigen promotes cell proliferation and tumorigenesis by up-regulating STAT3 in non-small cell lung cancer[J].Biomed Pharmacother,2018,104:595-602.DOI: 10.1016/j.biopha.2018.05.071. [29] ZhengW,XuS.Analysis of differential expression proteins of paclitaxel-treated lung adenocarcinoma cell A549 using tandem mass tag-based quantitative proteomics[J].Onco Targets Ther,2020,13:10297-10313.DOI: 10.2147/OTT.S259895. [30] ChiangCP,LangMJ,LiuBY,et al.Expression of proliferating cell nuclear antigen (PCNA) in oral submucous fibrosis, oral epithelial hyperkeratosis and oral epithelial dysplasia in Taiwan[J].Oral Oncol,2000,36(4):353-359.DOI: 10.1016/s1368-8375(00)00014-2. [31] ChenW,WuC,ChenY,et al.Downregulation of ceramide synthase 1 promotes oral cancer through endoplasmic reticulum stress[J].Int J Oral Sci,2021,13(1):10.DOI: 10.1038/s41368-021-00118-4. [32] KisK,LiuX,HagoodJS.Myofibroblast differentiation and survival in fibrotic disease[J].Expert Rev Mol Med,2011,13:e27.DOI: 10.1017/S1462399411001967. [33] ShenM,YoungA,C.PCNAAutexier, a focus on replication stress and the alternative lengthening of telomeres pathway[J].DNA Repair (Amst),2021,100:103055.DOI: 10.1016/j.dnarep.2021.103055. [34] DharadharS,van DijkWJ,ScheffersS,et al.Insert L1 is a central hub for allosteric regulation of USP1 activity[J].EMBO Rep,2021,22(4):e51749.DOI: 10.15252/embr.202051749. [35] LiB,WangR,WangY,et al.Regulation of smooth muscle contraction by monomeric non-RhoA GTPases[J].Br J Pharmacol,2020,177(17):3865-3877.DOI: 10.1111/bph.15172. [36] PfitzerG,WirthA,LuciusC,et al.Regulation of smooth muscle contraction by calcium, monomeric GTPases of the Rho subfamily and their effector kinases[J].Adv Exp Med Biol,2003,538:89-99; discussion 99.DOI: 10.1007/978-1-4419-9029-7_8. [37] LiX,WangF,LanY,et al.GDF-5 induces epidermal stem cell migration via RhoA-MMP9 signalling[J].J Cell Mol Med,2021,25(4):1939-1948.DOI: 10.1111/jcmm.15925. [38] QiY,LiangX,DaiF,et al.RhoA/ROCK pathway activation is regulated by AT1 receptor and participates in smooth muscle migration and dedifferentiation via promoting actin cytoskeleton polymerization[J].Int J Mol Sci,2020,21(15):5398. DOI: 10.3390/ijms21155398. [39] LeinhosL,PetersJ,KrullS,et al.Hypoxia suppresses myofibroblast differentiation by changing RhoA activity[J].J Cell Sci,2019,132(5):jcs223230. DOI: 10.1242/jcs.223230. [40] TsaiCH, LinBJ, ChaoPH. α2β1 integrin and RhoA mediates electric field-induced ligament fibroblast migration directionality[J].J Orthop Res, 2013,31(2):322-327. DOI: 10.1002/jor.22215. -

1 采用活细胞工作站观察电场刺激各时间点2组人皮肤成纤维细胞的形态和排列 倒置相差显微镜×100,图中标尺为100 μm。 1A、1B.分别为模拟电场组处理0(即刻)、6 h,细胞均呈任意方向排列;1C、1D.分别为200 mV/mm电场组处理0、6 h,图1C细胞呈任意方向排列,图1D细胞形态拉长,呈纵向排列,细胞长轴与电场方向垂直
注:图1D下方箭头为电场方向,“+” 为正极、“-” 为负极
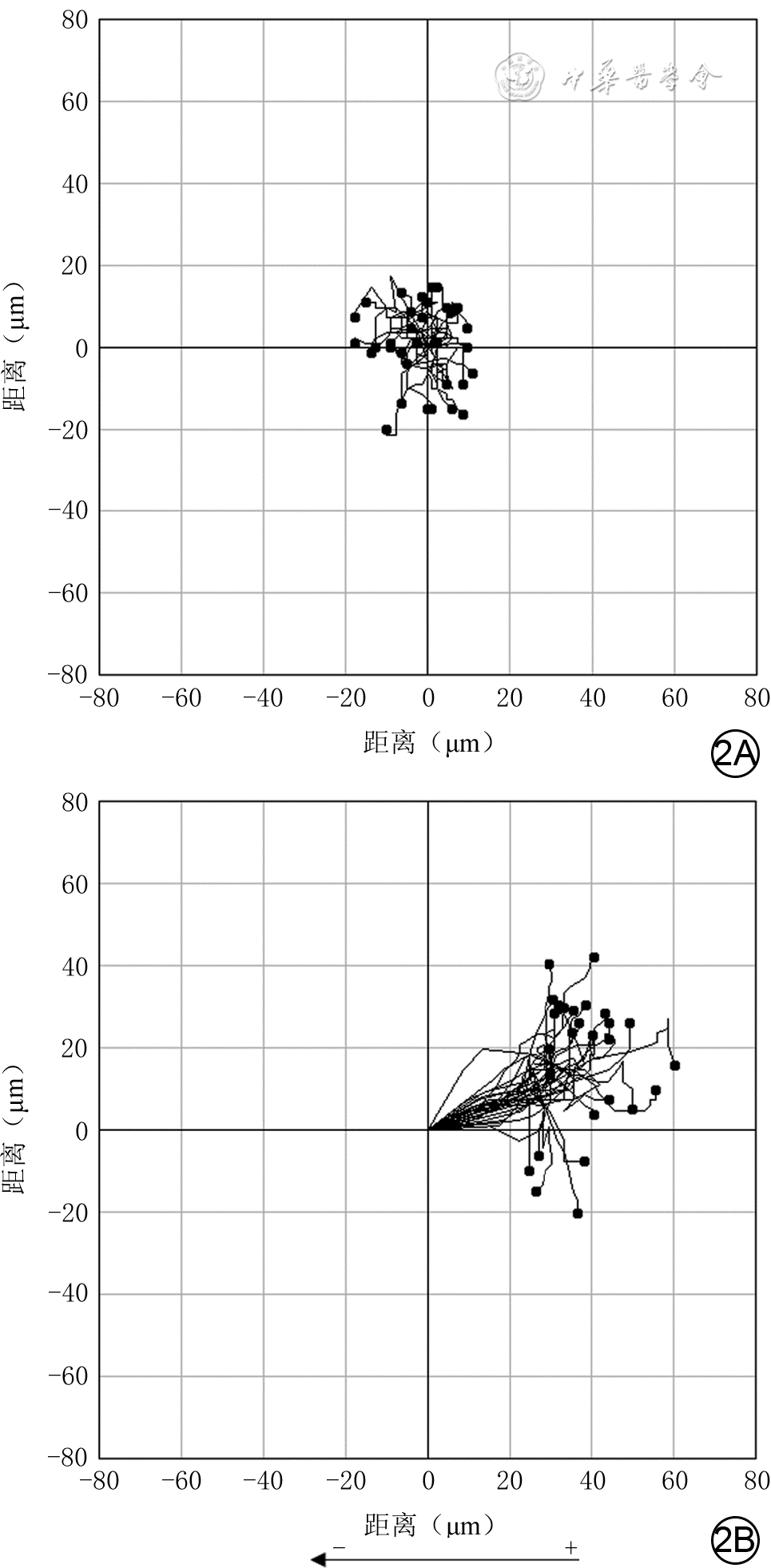
2 采用活细胞工作站观察2组人皮肤成纤维细胞经电场处理3 h内运动轨迹 倒置相差显微镜×100。2A.模拟电场组细胞绕原点运动;2B.200 mV/mm电场组细胞趋向于正极运动
注:细胞运动起点为坐标(0,0),运动终点为4个象限中的圆点,连接前述2个点之间的曲线为细胞运动轨迹,圆点位于左上、左下象限代表细胞向负极方向迁移,圆点位于右上、右下象限代表细胞向正极方向迁移;图2B下方箭头为电场方向,“ + ” 为正极、“-”为负极
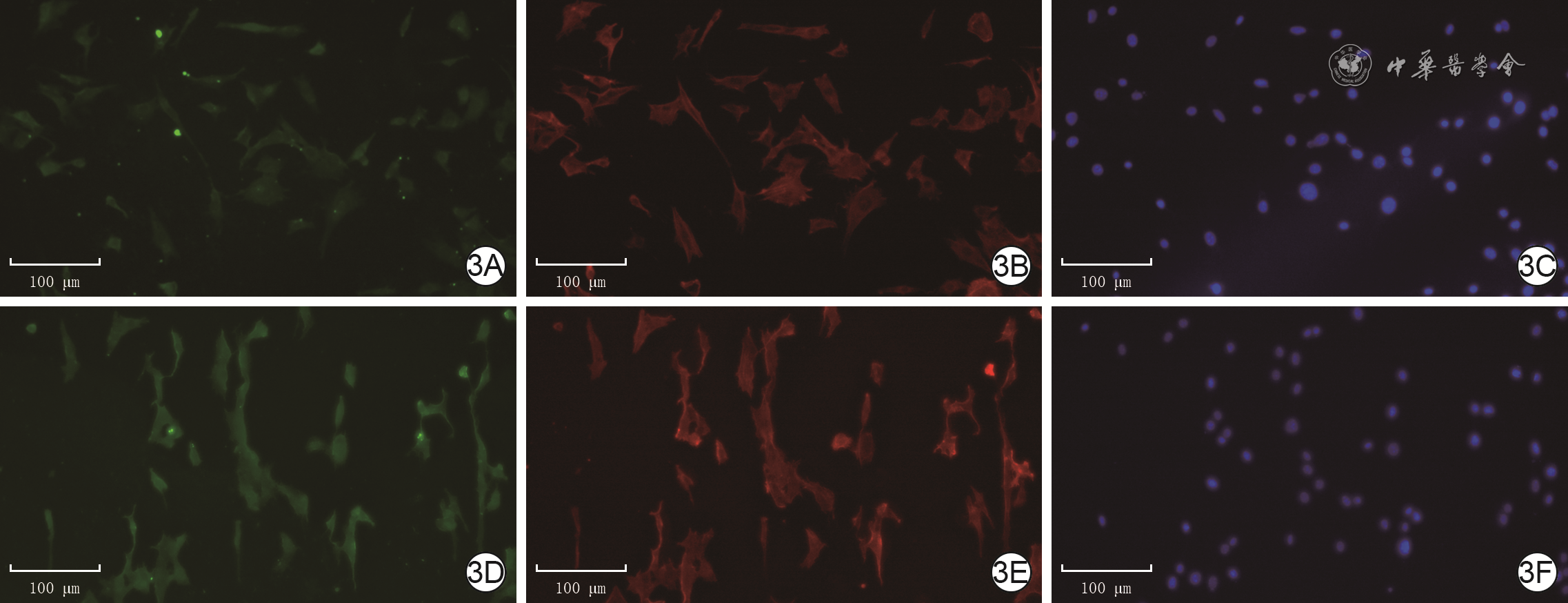
3 免疫荧光法检测电场处理3 h后2组人皮肤成纤维细胞α-SMA的表达 Alexa Fluor 488-4′,6-二脒基-2-苯基吲哚-罗丹明标记的鬼笔环肽×100,图中标尺为100 μm。3A、3B、3C.分别为模拟电场组细胞α-SMA蛋白表达、纤维状肌动蛋白排列、细胞核形态,α-SMA蛋白表达较低,纤维状肌动蛋白无序排列,细胞核完整;3D、3E、3F.分别为200 mV/mm电场组细胞α-SMA蛋白表达、纤维肌动蛋白排列、细胞核形态,图3D中α-SMA蛋白表达较图3A增加,纤维状肌动蛋白纵向排列,细胞核完整
注:绿色荧光标记α平滑肌肌动蛋白(α-SMA),红色荧光标记纤维状肌动蛋白,蓝色荧光标记细胞核
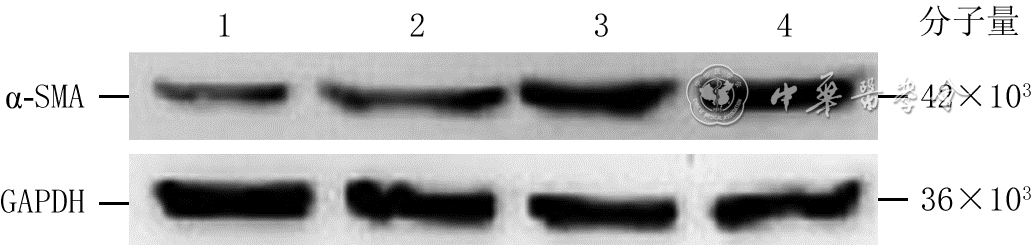
4 蛋白质印迹法检测不同强度电场处理3 h后4组人皮肤成纤维细胞α-SMA蛋白表达
注:α-SMA为α平滑肌肌动蛋白,GAPDH为3-磷酸甘油醛脱氢酶;条带上方1、2、3、4分别指模拟电场组、100 mV/mm电场组、200 mV/mm电场组、400 mV/mm电场组
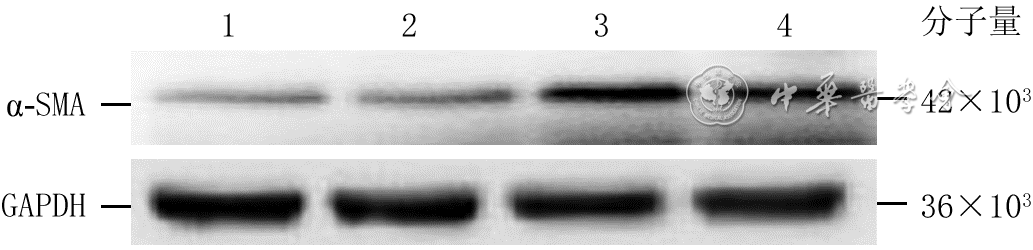
5 蛋白质印迹法检测电场处理不同时间4组人皮肤成纤维细胞α-SMA蛋白表达
注:α-SMA为α平滑肌肌动蛋白,GAPDH为3-磷酸甘油醛脱氢酶;条带上方1、2、3、4分别指模拟电场组(处理6 h)、电场处理1 h组、电场处理3 h组、电场处理6 h组

6 蛋白质印迹法检测不同强度电场处理3 h后4组人皮肤成纤维细胞增殖细胞核抗原(PCNA)蛋白表达
注:条带上方1、2、3、4分别指模拟电场组、100 mV/mm电场组、200 mV/mm电场组、400 mV/mm电场组
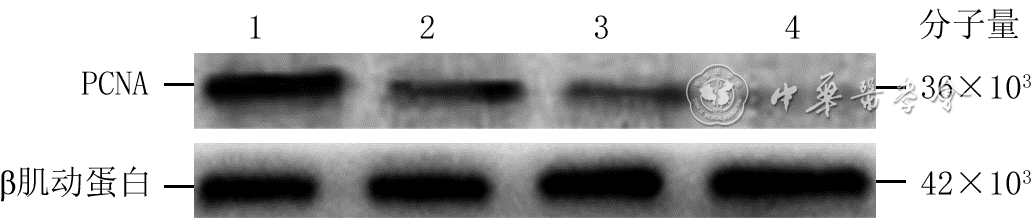
7 蛋白质印迹法检测电场处理不同时间4组人皮肤成纤维细胞增殖细胞核抗原(PCNA)蛋白表达
注:条带上方1、2、3、4分别指模拟电场组(处理6 h)、电场处理1 h组、电场处理3 h组、电场处理6 h组
表1 2组人皮肤成纤维细胞经电场处理3 h内cosθ及位移速度和轨迹速度比较[M(Q1,Q3)]
组别 样本数 cosθ 位移速度(μm/min) 轨迹速度(μm/min) 模拟电场组 34 -0.184(-0.336,0.371) 0.207(0.160,0.261) 0.165(0.126,0.204) 200 mV/mm电场组 30 0.833(0.694,0.925) 0.470(0.419,0.523) 0.383(0.347,0.438) Z值 -4.39 -5.33 -5.41 P值 <0.001 <0.001 <0.001  下载: 导出CSV
下载: 导出CSV
《中华烧伤与创面修复杂志》第六届编辑委员会通讯编委名单按姓氏拼音排序
贲道锋 卞徽宁 曹永倩 晁生武 陈辉 陈婧 陈朗 陈铭锐 陈鹏 陈晓东 陈忠勇 程君涛 迟云飞 储国平 党永明 邓呈亮 狄海萍 丁国兵 丁若虹 董茂龙 段红杰 段鹏 樊东力 房贺 冯光 付忠华 郭毅斌 韩兆峰 侯春胜 胡德林 胡炯宇 胡骁骅 胡晓燕 黄红军 纪世召 江华 姜丽萍 姜玉峰 雷娜 黎宁 李东杰 李峰 李靖 李晓东 李晓鲁 梁钢 梁鹏飞 林才 林国安 林源 刘德伍 刘健 刘军 刘淑华 龙奕 卢长虹 鲁峰 吕开阳 吕强 马思远 牛轶雯 欧阳军 乔亮 覃凤均 邱学文 曲滨 任超 沈江涌 石继红 宋慧锋 苏海涛 苏永涛 孙勇 孙瑜 谭江琳 唐修俊 滕苗 田社民 涂家金 汪虹 汪洋 王爱萍 王德怀 王洪涛 王会军 王良喜 王爽 王献珍 王志永 温冰 邬佳敏 吴红 吴继炎 吴巍巍 吴祖煌 向飞 向军 谢举临 谢松涛 辛海明 许喜生 许学文 薛斌 杨建民 杨敏烈 杨薛康 姚明 姚兴伟 叶祥柏 易成刚 易南 于东宁 岳丽青 翟红军 詹日兴 张博 张东霞 张红艳 张菊芳 张玲娟 张庆红 张彦琦 张寅 张元海 张志 赵全 赵冉 赵雄 郑德义 郑东风 郑军 周国富 周俊峄 周琴 周万芳 朱峰 朱宇刚 祝筱梅 邹立津 邹晓防
下载: 导出CSV
-


 下载:
下载:


计量
- 文章访问数: 422
- HTML全文浏览量: 89
- PDF下载量: 30
- 被引次数: 0
