



Biological activity of biomimetic dermal papilla spheres prepared by culture of dermal papilla cells of mice based on hanging drops of gelatin methacrylate and its hair-inducing function in nude mice
-
摘要:
目的 探讨基于甲基丙烯酸酐化明胶(GelMA)仿生微环境与悬滴法三维培养小鼠毛乳头细胞(DPC)制备仿生毛乳头球的生物活性及其在裸鼠中的毛发诱导功能。 方法 采用实验研究方法。采用酶消化法获取雄性5~6周龄C57BL/6J小鼠触须毛的DPC以及1 d龄C57BL/6J小鼠皮肤角质形成细胞(KC),经免疫荧光法鉴定前者第3代细胞稳定表达DPC标志物神经细胞黏附分子、碱性磷酸酶(ALP)、β连环蛋白及α平滑肌肌动蛋白,后者原代细胞稳定表达KC标志物角蛋白15。取第8代DPC,用GelMA重新悬浮并接种至Transwell孔板插件底面,光交联后倒置培养,于光学显微镜下观察培养0(即刻)、3 d GelMA悬滴中DPC聚集情况(聚集成球即仿生毛乳头球),采用活/死染色试剂盒检测培养3 d仿生毛乳头球中细胞活性。将传统二维培养的原代DPC、第8代DPC以及按前述方法制备的仿生毛乳头球分别设为原代DPC组、第8代DPC组、仿生毛乳头球组,利用高通量测序技术平台对每组培养3 d的3个样本中DPC进行转录组测序,利用基迪奥生物信息云工具平台(OmicShare Tools)对转录组数据进行主成分分析、Pearson相似性分析、差异表达基因筛选,利用时间序列趋势分析软件对差异表达基因进行表达模式聚类分析,利用基迪奥生物信息云工具平台对具有特定表达模式的差异表达基因进行京都基因与基因组百科全书(KEGG)和基因本体论(GO)富集分析。同前进行细胞分组,采用随机数字表法从差异表达基因中抽取性别决定区Y框蛋白8(SOX8)、基质金属蛋白酶9(MMP-9)、26型胶原纤维α1(COL26A1)、无翅型MMTV整合位点家族成员6(Wnt6),采用实时荧光定量反转录PCR(RT-PCR)法验证差异表达基因mRNA表达情况与测序结果的一致性(样本数为9);采用实时荧光定量RT-PCR法检测DPC生物功能标志物成纤维细胞生长因子7(FGF7)、Wnt10a、淋巴样增强因子1(LEF1)、ALP、β连环蛋白、多功能蛋白聚糖、SOX2的mRNA表达情况(样本数为9)。取3只5~6周龄雄性BALB/c裸鼠,分别设为原代DPC组、第8代DPC组、仿生毛乳头球组,将原代DPC、第8代DPC、仿生毛乳头球分别与原代KC以2∶1细胞数比例混合后分别注射至相应组别裸鼠皮下,每只注射6个区域。注射后2周,取注射区域全厚皮,计数再生毛发,行苏木精-伊红染色后观察再生毛囊情况。对数据行单因素方差分析、Tukey检验并进行Bonferroni校正。 结果 培养3 d,DPC在GelMA悬滴中由培养0 d的分散状态聚集成仿生毛乳头球,制备的仿生毛乳头球中细胞活性良好。培养3 d,主成分分析显示,相比于第8代DPC组,原代DPC组、仿生毛乳头球组组内样本间变异程度较低,仿生毛乳头球组与原代DPC组组间样本变异程度最低,3组DPC样本超过90%的基因谱数据变异可由第1个和第2个主要成分来解释;Pearson相似性分析显示,组内样本重复性好,原代DPC组和仿生毛乳头球组样本间的相关系数为0.84~0.95,原代DPC组和第8代DPC组样本间的相关系数为0.72~0.87;3组DPC间差异表达基因分析筛选出642个组间交集差异表达基因,对其进行表达模式聚类显示有2种基因表达模式有显著趋势(P<0.05),分别为第8代DPC组基因表达显著低于原代DPC组/仿生毛乳头球组、第8代DPC组基因表达显著高于原代DPC组/仿生毛乳头球组,共包含411个差异表达基因;KEGG富集分析显示这411个差异表达基因在Wnt信号通路以及磷脂酰肌醇3激酶-蛋白激酶B信号通路显著富集(P<0.05),GO富集分析表明细胞外基质、经典Wnt信号通路、细胞分化等GO术语被显著富集(P<0.05)。与原代DPC组、仿生毛乳头球组比较,第8代DPC组细胞基因SOX8、MMP-9、COL26A1、Wnt6 mRNA表达量均明显下降(q=15.950、8.854、11.890、11.050,9.851、5.884、7.418、4.870,P<0.01),与测序数据一致。与原代DPC组、仿生毛乳头球组比较,第8代DPC组细胞生物功能标志物FGF7、Wnt10a、LEF1、ALP、β连环蛋白、多功能蛋白聚糖、SOX2 mRNA表达量均明显下降(q=11.470、9.795、4.165、9.242、10.970、10.570、8.005,7.472、4.976、3.651、4.784、5.236、6.825、5.214,P<0.05或P<0.01)。注射后2周,第8代DPC组裸鼠无毛发再生,仿生毛乳头球组与原代DPC组裸鼠再生毛发数量相近(q=1.852,P>0.05)且均显著高于第8代DPC组(q=18.980、17.130,P<0.01);第8代DPC组裸鼠注射区域皮肤仅形成了坏死灶,而仿生毛乳头球组与原代DPC组裸鼠注射区域皮肤均观察到再生毛囊且毛囊横断切面有黑色素沉着。 结论 基于GelMA仿生微环境与悬滴法三维培养小鼠DPC制备仿生毛乳头球的培养模型可一定程度上恢复高传代DPC在裸鼠中的毛发诱导能力,且其生物特性更加类似于原代DPC,可实现DPC的扩增后特性恢复。 Abstract:Objective To investigate the biological activity of biomimetic dermal papilla spheres (DPSs) prepared by three-dimensionally cultured dermal papilla cells (DPCs) of mice based on the biomimetic microenvironment of gelatin methacrylate (GelMA) and hanging drop method and its hair-inducing function in nude mice. Methods Experimental research methods were adopted. DPCs from the vibrissa of male C57BL/6J mice aged 5 to 6 weeks and keratinocytes (KCs) from the skin of 1 d old C57BL/6J mice were obtained by enzymatic digestion method. A stable expression of DPCs markers such as nerve cell adhesion molecules, alkaline phosphatase (ALP), β-catenin, and α-smooth muscle actin were identified by immunofluorescence method in the third passage of the former cells, while the latter primary cells stably expressed keratin 15, a marker of KCs. The 8th passage of DPCs were re-suspended with GelMA and inoculated on the bottom surface of the Transwell plate insert, and then the GelMA drops were photocrosslinked and cultured upside down later. The DPCs aggregation in GelMA drops after in cultures of 0 (immediately) and 3 day (s) was observed under an optical microscope (the DPCs aggregates were the biomimetic DPSs). The cell viability of 3 day biomimetic DPSs culture was detected by live/dead staining kit. The primary DPCs and the 8th passage of DPCs derived from traditionally two-dimensional cultures, and the biomimetic DPSs prepared by the above-mentioned method were set as primary DPCs group, the 8th passage of DPCs group, and biomimetic DPSs group, respectively. Transcriptome sequencing was performed using the high-throughput sequencing technology platform, with 3 samples in each group analyzed after three days in culture. The principal component analysis, Pearson similarity analysis, and screening of differentially expressed genes were performed using OmicShare Tools based on the transcriptome data. Cluster analysis of expression patterns of differentially expressed genes was performed using time series trend analysis software. The Kyoto Encyclopedia of Genes and Genomes (KEGG) and Gene Ontology (GO) enrichment analyses of differentially expressed genes with specific expression patterns were performed using the OmicShare Tools. The cells were grouped as before, and the sex determining region Y-Box 8 (SOX8), matrix metallopeptidase 9 (MMP-9), collagen type ΧΧⅥ alpha 1 chain (COL26A1), and wingless-type MMTV integration site family member 6 (Wnt6) were screened out from the differentially expressed genes according to the random number table, which were determined by real-time fluorescent quantitative reverse transcription polymerase chain reaction (RT-PCR) to verify the consistency between mRNA expression of differentially expressed genes and sequencing results (n=9); the mRNA expressions of DPCs biological function markers fibroblast growth factor 7 (FGF7), Wnt10a, lymphoid enhancement factor 1 (LEF1), ALP, β-catenin, versican, and SOX2 were determined by real-time fluorescent quantitative RT-PCR (n=9). Three male BALB/c nude mice aged 5-6 weeks were divided into primary DPCs group, the 8th passage of DPCs group, and biomimetic DPSs group. The primary DPCs, the 8th passage of DPCs, and the biomimetic DPSs were mixed with primary KCs at a ratio of 2∶1 in cell number and then injected subcutaneously into mice of corresponding groups, with 6 injection regions for each mouse. Two weeks after the injection, the full-thickness skin of the injection region was taken, the regenerated hair was counted, and the regenerated hair follicle was observed after hematoxylin-eosin staining. Data were statistically analyzed with one-way analysis of variance, Tukey test, and Bonferroni correction. Results After 3 days of culture, DPCs aggregated into biomimetic DPSs in GelMA hanging drops from the dispersed state on culture day 0, and the cells in the biomimetic DPSs had good cell activity. After 3 days of culture, principal component analysis showed that compared with that of the 8th passage of DPCs group, the degree of variation among samples within primary DPCs group and biomimetic DPSs group was relatively low, and the degree of variation was the lowest between samples of biomimetic DPSs group and primary DPCs group; the variation of gene profile data of more than 90% of DPCs samples in 3 groups could be explained by the first and second principal components. Pearson similarity analysis showed that samples within each group had good repeatability. The similarity was 0.84-0.95 between samples of primary DPCs group and biomimetic DPSs group, and was 0.72-0.87 between samples of primary DPCs group and the 8th passage of DPCs group. The differentially expressed genes among the three groups were analyzed and 642 differentially expressed genes with group intersection were screened out. Clusters of expression patterns showed that two gene expression patterns had a significant trend (P<0.05), the first pattern showed that gene expression of the 8th passage of DPCs group was significantly lower than that of primary DPCs group or biomimetic DPSs group, and the second pattern showed that gene expression of the 8th passage of DPCs group was significantly higher than that of primary DPCs group or biomimetic DPSs group, including a total of 411 differentially expressed genes. KEGG enrichment analysis showed that the 411 differentially expressed genes were significantly enriched in Wnt signaling pathway and phosphatidylinositol 3 kinase/protein kinase B pathway (P<0.05), while GO enrichment analysis showed that GO terms such as extracellular matrix, classical Wnt signaling pathway, and cell differentiation were significantly enriched (P<0.05). The mRNA expressions of genes SOX8, MMP-9, COL26A1 and Wnt6 of cells in the 8th passage of DPCs group were significantly decreased compared with those in primary DPCs group and biomimetic DPSs group (q=15.950, 8.854, 11.890, 11.050, 9.851, 5.884, 7.418, 4.870, P<0.01), consistent with the sequencing data. Compared with those in primary DPCs group and biomimetic DPSs group, the mRNA expressions of biological function markers FGF7, Wnt10a, LEF1, ALP, β-catenin, versican, and SOX2 of cells in the 8th passage of DPCs group were significantly decreased (q=11.470, 9.795, 4.165, 9.242, 10.970, 10.570, 8.005, 7.472, 4.976, 3.651, 4.784, 5.236, 6.825, 5.214, P<0.05 or P<0.01). Two weeks after injection, nude mice in the 8th passage of DPCs group failed to regenerate hair, while the numbers of hair regenerated in nude mice in biomimetic DPSs group and primary DPCs group were close (q=1.852, P>0.05) and both were significantly higher than the number in the 8th passage of DPCs group (q=18.980, 17.130, P<0.01). In the 8th passage of DPCs group, only necrotic foci were found in the injection region of the skin of nude mice, while regenerated hair follicles were observed in the injection region of the skin of nude mice in both biomimetic DPSs group and primary DPCs group, and melanosis was observed in the cross section of hair follicles. Conclusions Based on GelMA biomimetic microenvironment and hanging drop method, the biomimetic DPSs culturing model prepared by three dimensional culture of DPCs of mice can restore the hair-inducing ability of high passage of DPCs in nude mice to a certain extent, and its biological characteristics are more similar to those of the primary DPCs, which can restore the characteristics of DPCs after amplification. -
参考文献
(36) [1] 唐建兵,李勤,程飚,等.毛乳头在毛囊生长周期中的变化和作用[J].南方医科大学学报,2008,28(9):1649-1651.DOI: 10.3321/j.issn:1673-4254.2008.09.046. [2] HouschyarKS,BorrelliMR,TapkingC,et al.Molecular mechanisms of hair growth and regeneration: current understanding and novel paradigms[J].Dermatology,2020,236(4):271-280.DOI: 10.1159/000506155. [3] 刘公言,白莉雅,李福昌,等.毛囊发育与周期性生长的调控信号通路研究进展[J]. 畜牧与兽医,2021,53(1):125-129. [4] GentileP,GarcovichS.Advances in regenerative stem cell therapy in androgenic alopecia and hair loss: wnt pathway, growth-factor, and mesenchymal stem cell signaling impact analysis on cell growth and hair follicle development[J].Cells,2019,8(5):466.DOI: 10.3390/cells8050466. [5] 胡天星,杨希川.毛乳头分子特征研究进展[J].临床皮肤科杂志,2020,49(3):187-189.DOI: 10.16761/j.cnki.1000-4963.2020.03.017. [6] 黄丽结,祝宁侠,谢小琴,等. 毛乳头细胞诱导毛囊再生的研究现状[J]. 科学咨询,2019(42):120-121. [7] WangB,LiuXM,LiuZN,et al.Human hair follicle-derived mesenchymal stem cells: isolation, expansion, and differentiation[J].World J Stem Cells,2020,12(6):462-470.DOI: 10.4252/wjsc.v12.i6.462. [8] TaghiabadiE,NilforoushzadehMA,AghdamiN.Maintaining hair inductivity in human dermal papilla cells: a review of effective methods[J].Skin Pharmacol Physiol,2020,33(5):280-292.DOI: 10.1159/000510152. [9] SaxenaN,MokKW,RendlM.An updated classification of hair follicle morphogenesis[J].Exp Dermatol,2019,28(4):332-344.DOI: 10.1111/exd.13913. [10] 伍津津.毛囊干细胞与组织工程研究进展[J].中国医学文摘(皮肤科学),2016,33(4):415-423. [11] FotyR. A simple hanging drop cell culture protocol for generation of 3D spheroids[J]. J Vis Exp,2011(51):2720. DOI: 10.3791/2720. [12] 邓俊豪,李苗,张里程,等.三维悬滴法培养间充质干细胞在组织损伤修复中的应用及优势[J].中国组织工程研究,2020,24(7):1101-1106.DOI: 10.3969/j.issn.2095-4344.1849. [13] LinB,MiaoY,WangJ,et al.Surface tension guided hanging-drop: producing controllable 3D spheroid of high-passaged human dermal papilla cells and forming inductive microtissues for hair-follicle regeneration[J].ACS Appl Mater Interfaces,2016,8(9):5906-5916.DOI: 10.1021/acsami.6b00202. [14] LiuBC,WangY,MiaoY,et al.Hydrogen bonds autonomously powered gelatin methacrylate hydrogels with super-elasticity, self-heal and underwater self-adhesion for sutureless skin and stomach surgery and E-skin[J].Biomaterials,2018,171:83-96.DOI: 10.1016/j.biomaterials.2018.04.023. [15] Dursun UsalT,YucelD,HasirciV.A novel GelMA-pHEMA hydrogel nerve guide for the treatment of peripheral nerve damages[J].Int J Biol Macromol,2019,121:699-706.DOI: 10.1016/j.ijbiomac.2018.10.060. [16] 王毅虎,曹霄峰,张兵,等.甲基丙烯酰胺基明胶水凝胶研究进展[J].影像科学与光化学,2020,38(1):22-26.DOI: 10.7517/issn.1674-0475.190612. [17] 陈琳,习佳飞,刘大庆,等.小鼠皮肤表皮和真皮细胞共移植诱导毛囊再生实验研究[J].中国修复重建外科杂志,2016,30(4):485-490.DOI: 10.7507/1002-1892.20160097. [18] MiJX,ZhangYN,LaiZH,et al.Principal component analysis based on nuclear norm minimization[J].Neural Netw,2019,118:1-16.DOI: 10.1016/j.neunet.2019.05.020. [19] ErnstJ,Bar-JosephZ.STEM: a tool for the analysis of short time series gene expression data[J].BMC Bioinformatics,2006,7:191.DOI: 10.1186/1471-2105-7-191. [20] ZhaoYW,WangJ,ChenJ,et al.A literature review of gene function prediction by modeling gene ontology[J].Front Genet,2020,11:400.DOI: 10.3389/fgene.2020.00400. [21] KanehisaM,FurumichiM,TanabeM,et al.KEGG: new perspectives on genomes, pathways, diseases and drugs[J].Nucleic Acids Res,2017,45(D1):D353-D361.DOI: 10.1093/nar/gkw1092. [22] 于秀菊,范阔海,贺俊平,等.小鼠自然毛囊生长周期及β连环蛋白的表达[J].解剖学报,2010,41(6):862-866.DOI: 10.3969/j.issn.0529-1356.2010.06.017. [23] 陈明娟,胡志奇,谭挺. 新生鼠真皮细胞的分离培养及其毛囊诱导能力的鉴定[J]. 中国美容整形外科杂志,2010,21(6):382-384. DOI: 10.3969/j.issn.1673-7040.2010.06.026. [24] HigginsCA,ChenJC,CeriseJE,et al.Microenvironmental reprogramming by three-dimensional culture enables dermal papilla cells to induce de novo human hair-follicle growth[J].Proc Natl Acad Sci U S A,2013,110(49):19679-19688.DOI: 10.1073/pnas.1309970110. [25] BetriuN, Jarrosson-MoralC, SeminoCE. Culture and differentiation of human hair follicle dermal papilla cells in a soft 3D self-assembling peptide scaffold[J]. Biomolecules,2020,10(5):684. DOI: 10.3390/biom10050684. [26] TopouziH,LoganNJ,WilliamsG,et al.Methods for the isolation and 3D culture of dermal papilla cells from human hair follicles[J].Exp Dermatol,2017,26(6):491-496.DOI: 10.1111/exd.13368. [27] 林鹤,王婉秋,焦佳媛,等.培养板悬滴法三维细胞培养模型建立及细胞活力检测方法比较[J].药物评价研究,2017,40(8):1103-1106.DOI: 10.7501/j.issn.1674-6376.2017.08.013. [28] GuptaAC,ChawlaS,HegdeA,et al.Establishment of an in vitro organoid model of dermal papilla of human hair follicle[J].J Cell Physiol,2018,233(11):9015-9030.DOI: 10.1002/jcp.26853. [29] WangJ,MiaoY,HuangY,et al.Bottom-up nanoencapsulation from single cells to tunable and scalable cellular spheroids for hair follicle regeneration[J].Adv Healthc Mater,2018,7(3).DOI: 10.1002/adhm.201700447. [30] 刘芳,苗勇,胡志奇. 毛乳头细胞生物学特性的研究进展[J]. 中华整形外科杂志,2021,37(5):567-570. DOI: 10.3760/cma.j.cn114453-20210408-00156. [31] 郭晓静,郝飞,姜晓勇,等.人毛乳头细胞在不同传代时期某些细胞因子表达的变化[J].第三军医大学学报,2006,28(21):2144-2145.DOI: 10.3321/j.issn:1000-5404.2006.21.010. [32] 蔡博治,陈先才,倪娜,等.鼠胚胎期和出生后毛囊下段启动再生表达的Wnt5a[J].中国组织工程研究,2020,24(31):5018-5022.DOI: 10.3969/j.issn.2095-4344.2136. [33] 田宇,宋志强,郝飞,等.体外凝集性生长状态下毛乳头细胞特异表达基因分析[J].中国实用医药,2009,4(29):27-29.DOI: 10.3969/j.issn.1673-7555.2009.29.013. [34] 郝建春,刘玮,袁小英,等.骨形成蛋白-2、成纤维细胞生长因子-2对大鼠触须毛乳头细胞增殖和毛发诱导能力的影响[J/CD].中华临床医师杂志:电子版,2015,9(11):2149-2153.DOI: 10.3877/cma.j.issn.1674-0785.2015.11.028. [35] VarkeyM,VisscherDO,van ZuijlenP,et al.Skin bioprinting: the future of burn wound reconstruction?[J/OL].Burns Trauma,2019,7:4[2021-03-30]. https://pubmed.ncbi.nlm.nih.gov/30805375/.DOI: 10.1186/s41038-019-0142-7. [36] GreenhalghDG.Management of facial burns[J/OL].Burns Trauma,2020,8:tkaa023[2021-03-30].https://academic.oup.com/burnstrauma/article/doi/ 10.1093/burnst/tkaa023/5867665.DOI: 10.1093/burnst/tkaa023. -

1 于BALB/c裸鼠真皮层注射成年小鼠第8代毛乳头细胞与新生小鼠角质形成细胞混悬液(细胞数比例2∶1)诱导毛囊再生
注:图中红色圆圈直径1 cm,为注射前皮丘范围标记,防止皮丘越过划定范围引起邻近注射点皮丘融合

2 免疫荧光染色法检测成年小鼠第3代毛乳头细胞(DPC)培养9 d以及新生小鼠原代角质形成细胞(KC)培养2 d相应细胞标志物表达 Alexa Fluor 594-Alexa Fluor 488-4',6-二脒基-2-苯基吲哚×50,图中标尺为100 μm。2A.DPC稳定表达神经细胞黏附分子(红色);2B.DPC稳定表达碱性磷酸酶(绿色)、β连环蛋白(红色);2C.DPC稳定表达α平滑肌肌动蛋白(绿色);2D.KC稳定表达角蛋白15(红色)

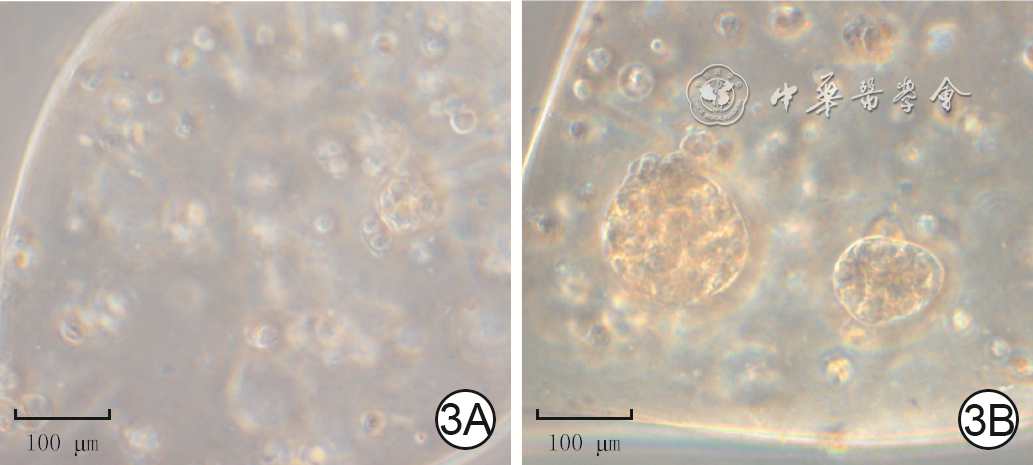
3 培养0、3 d甲基丙烯酸酐化明胶悬滴中小鼠毛乳头细胞(DPC)聚集情况 光学显微镜×20,图中标尺为100 μm。3A.培养0 d,DPC呈分散状态;3B.培养3 d,DPC聚集成球状,即仿生毛乳头球

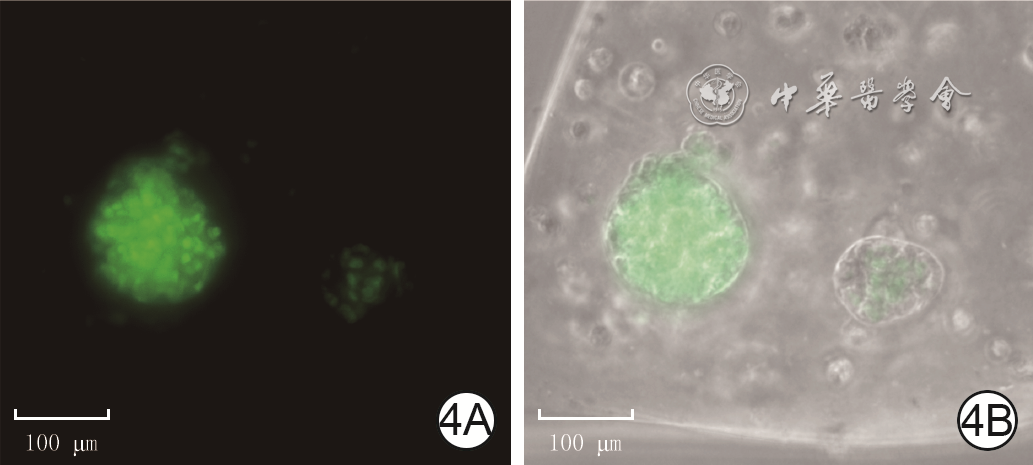
4 培养3 d制备的仿生毛乳头球中小鼠毛乳头细胞的活性 Alexa Fluor 488×20,图中标尺为100 μm。4A.细胞活性良好;4B.中心区域未见细胞坏死
注:活细胞染绿色,死细胞染红色

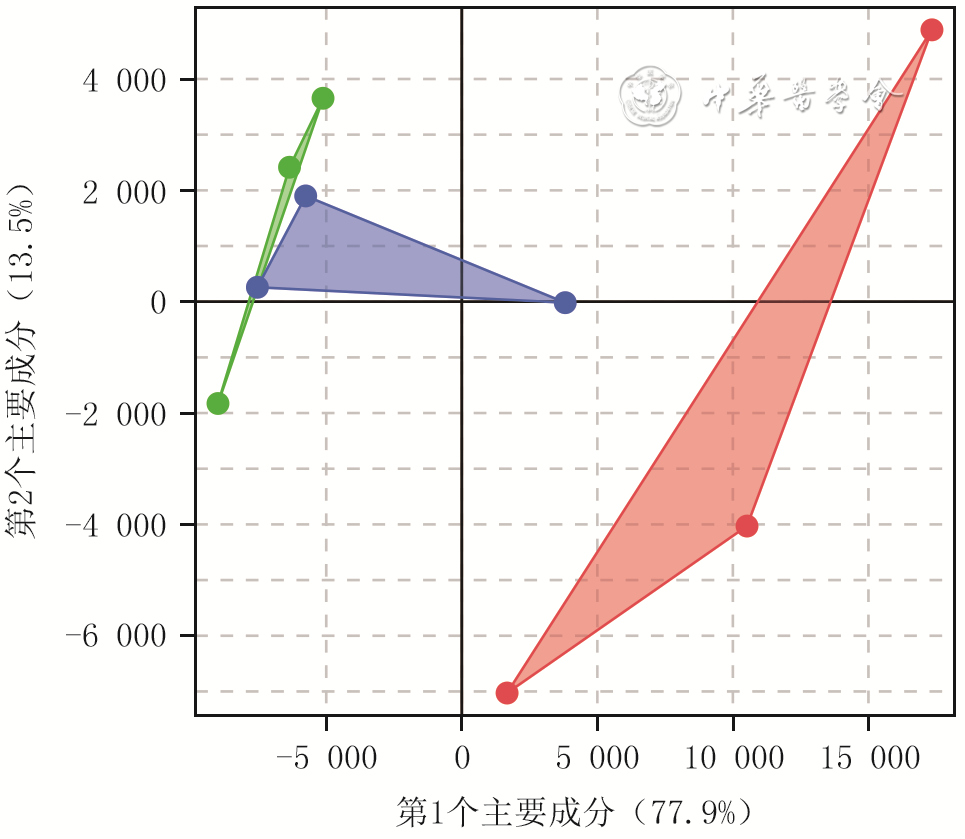
5 3组小鼠毛乳头细胞(DPC)培养3 d测序样本转录表达谱主成分分析(样本数为3)
注:绿色、红色、蓝色三角分别代表原代DPC组、第8代DPC组、仿生毛乳头球组样本表达谱数据降维后的二维投影

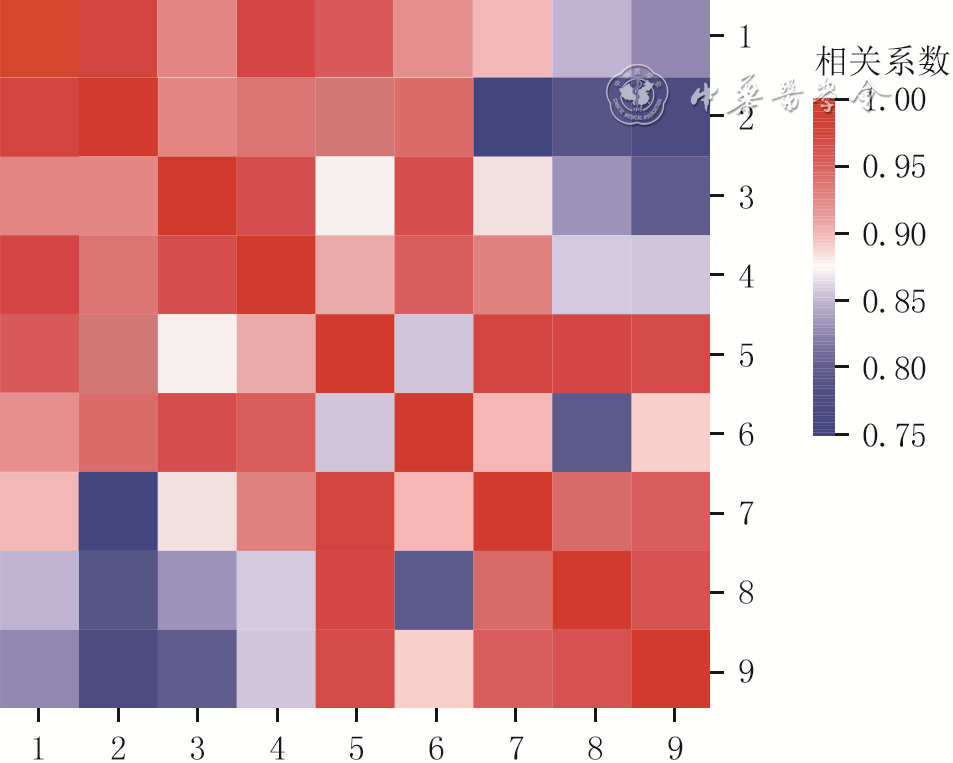
6 3组小鼠毛乳头细胞(DPC)培养3 d测序样本转录表达谱Pearson相似性分析(样本数为3)
注:矩阵下方和右侧1~3、4~6、7~9分别为原代DPC组、仿生毛乳头球组、第8代DPC组的3个样本

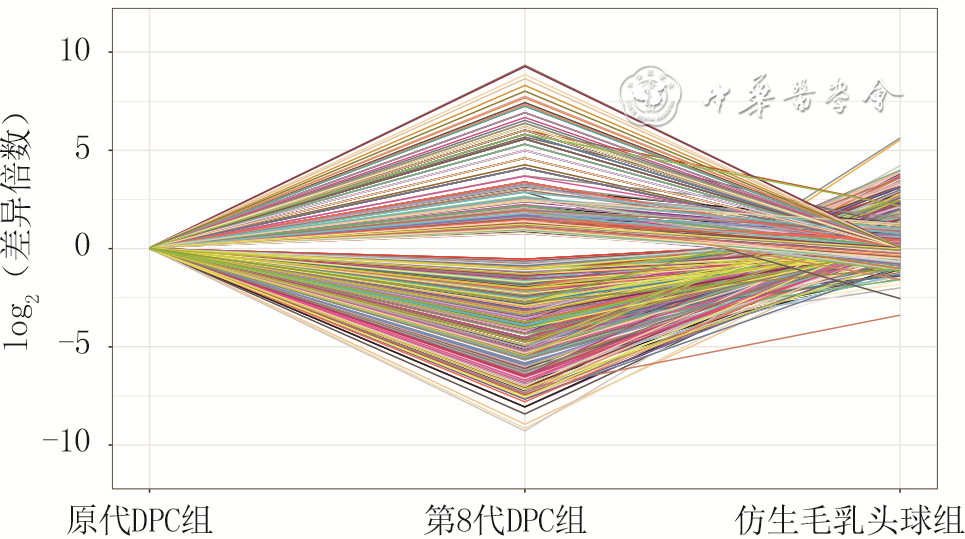
7 3组小鼠毛乳头细胞(DPC)培养3 d测序样本间642个交集差异表达基因表达模式聚类分析得出411个具特色表达模式的差异表达基因(样本数为3)
注:差异倍数(fold change)为第8代DPC组或仿生毛乳头球组与原代DPC组基因表达量的比值,图中每条线反映1个基因的表达变化程度,>0为上调,<0为下调

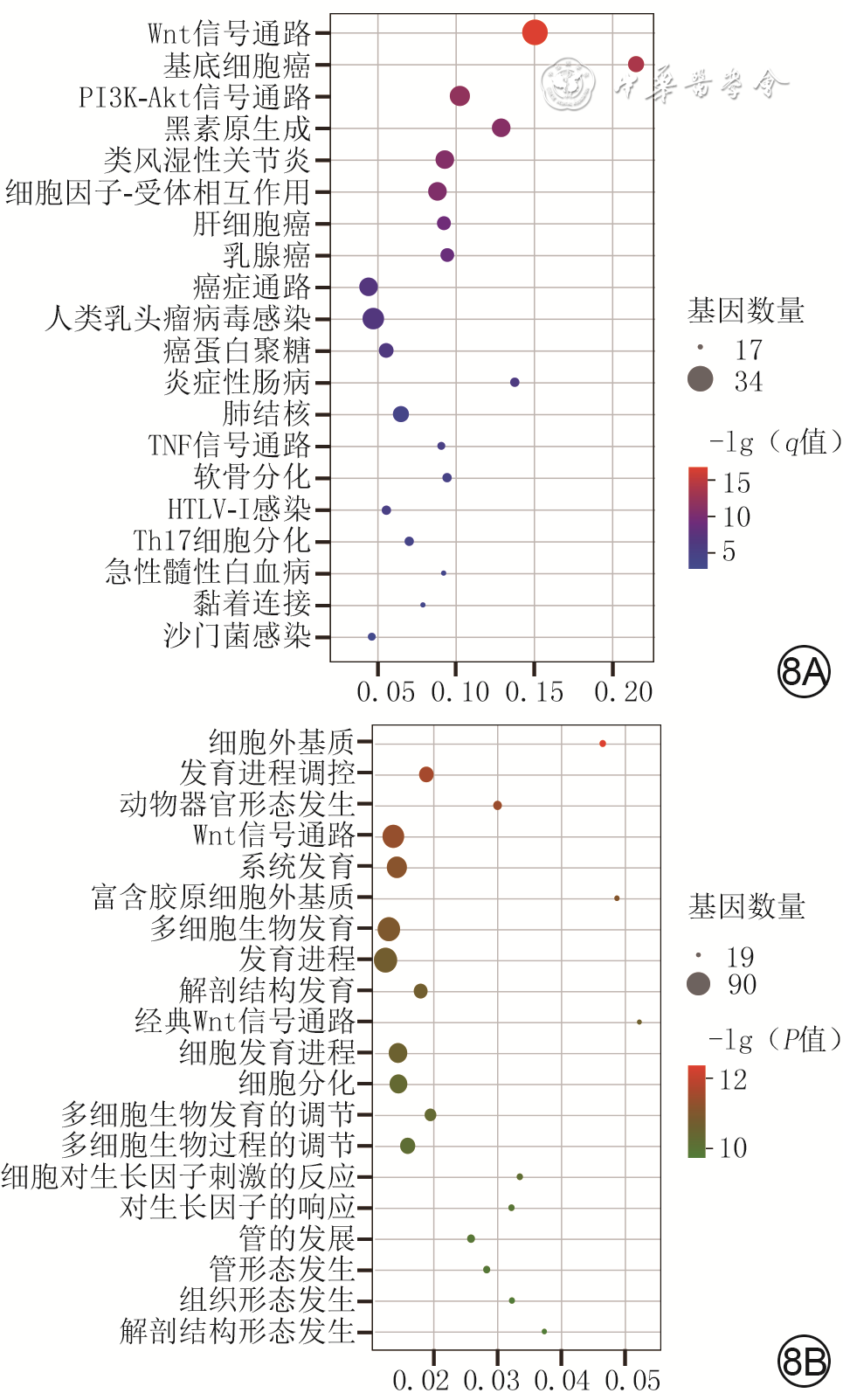
8 3组小鼠毛乳头细胞培养3 d测序样本间符合特定表达模式的411个差异表达基因的京都基因与基因组百科全书(KEGG)和基因本体论(GO)富集分析。 8A.富集程度排名前20差异表达基因的KEGG通路富集图;8B.富集程度排名前20差异表达基因的GO富集分析图
注:Wnt为无翅型MMTV整合位点家族成员,PI3K-Akt为磷脂酰肌醇3激酶-蛋白激酶B,TNF为肿瘤坏死因子,HTLV为嗜T细胞病毒,Th为辅助性T淋巴细胞;纵坐标为富集到的KEGG通路或GO术语,横坐标为富集分数(反映富集集中度);气泡大小反映对应KEGG通路或GO术语富集到的差异表达基因数量;q值、P值代表富集程度

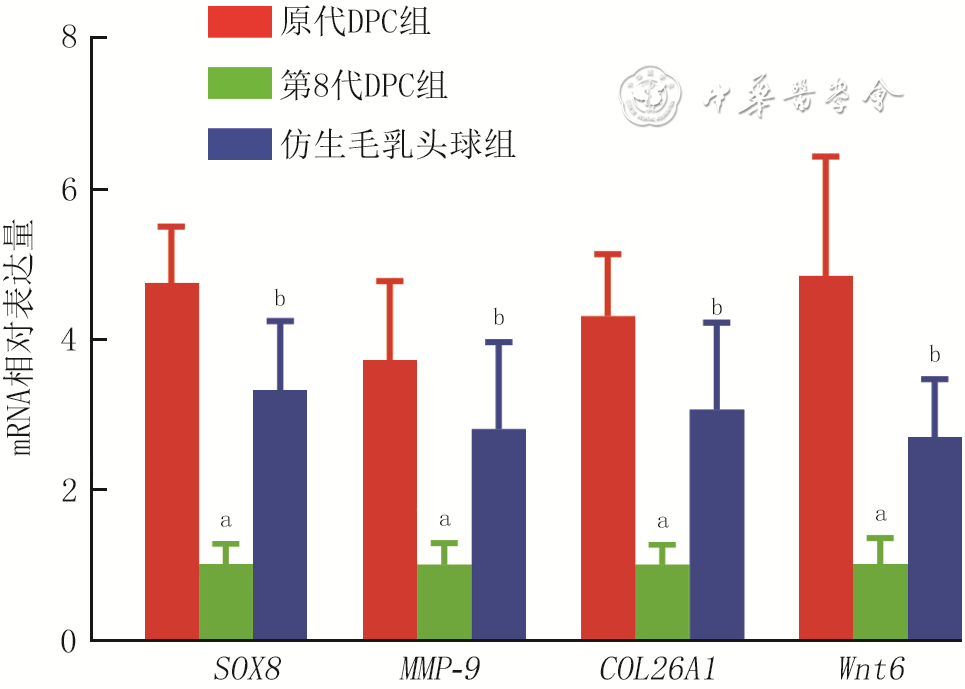
9 实时荧光定量反转录PCR法检测3组小鼠毛乳头细胞(DPC)培养3 d具有显著趋势的基因表达模式的基因性别决定区Y框蛋白8(SOX8)、基质金属蛋白酶9(MMP-9)、26型胶原纤维α1(COL26A1)、无翅型MMTV整合位点家族成员6(Wnt6)mRNA表达(样本数为9,
注:与原代DPC组比较,aP<0.01;与第8代DPC组比较,bP<0.01

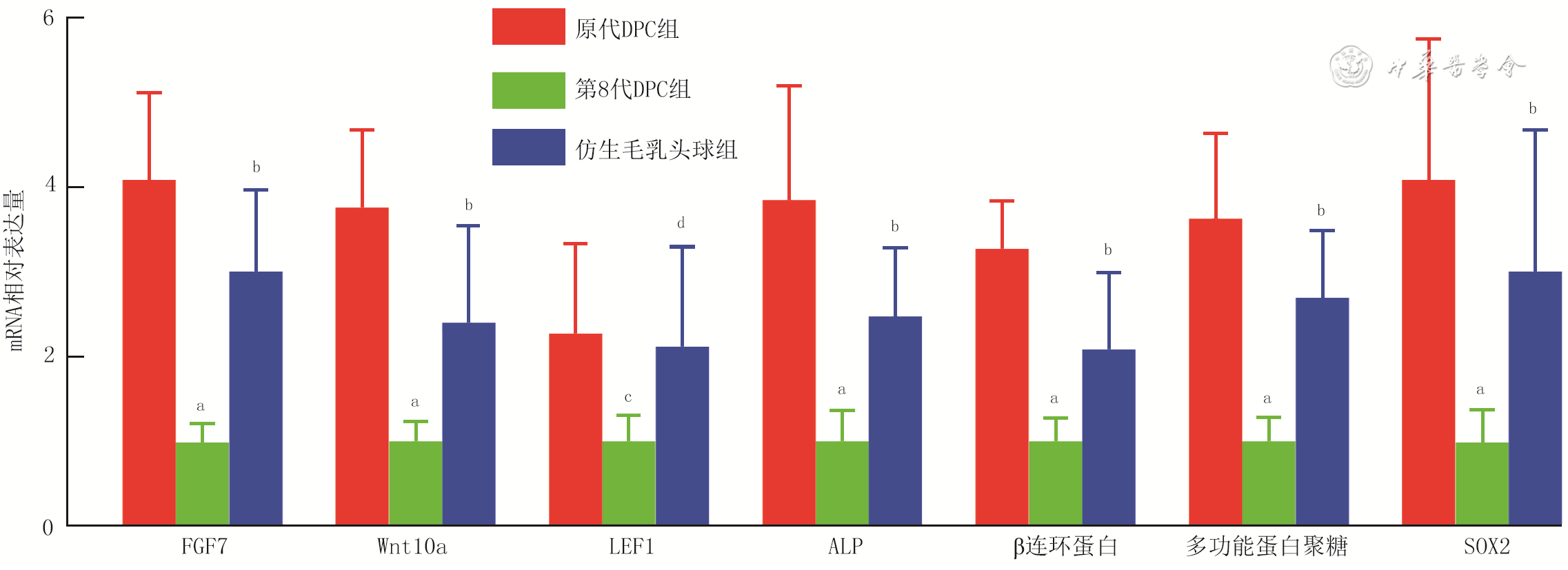
10 实时荧光定量反转录PCR法检测3组小鼠毛乳头细胞(DPC)培养3 d成纤维细胞生长因子7(FGF7)、无翅型MMTV整合位点家族成员10a(Wnt10a)、淋巴样增强因子1(LEF1)、碱性磷酸酶(ALP)、β连环蛋白、多功能蛋白聚糖、性别决定区Y框蛋白2(SOX2)mRNA表达(样本数为9,
注:与原代DPC组比较,aP<0.01,cP<0.05;与第8代DPC组比较,bP<0.01,dP<0.05

11 3组裸鼠皮肤注射成年小鼠毛乳头细胞(DPC)与新生小鼠角质形成细胞混悬液(细胞数比例2∶1)2周后注射区域再生毛发与再生毛囊情况。11A、11B、11C.分别为原代DPC组、仿生毛乳头球组、第8代DPC组再生毛发情况,图11A与图11B再生毛发数相近,图11C未见新生毛发生长 体视镜×50,图中标尺为500 μm;11D、11E、11F.分别为原代DPC组、仿生毛乳头球组、第8代DPC组再生毛囊情况,图11D与图11E可见新生毛囊(➝),图11F可见坏死灶(➝) 苏木精-伊红×200,图中标尺为50 μm
表1 实时荧光定量反转录PCR法检测毛乳头细胞差异表达基因mRNA表达的引物序列及产物大小
引物名称 引物序列(5'→3') 产物大小(bp) 性别决定区Y框蛋白8 上游:CTGGGACCAATGGATGCTGT 495 下游:TCAATGCCTGGCCCAATGAT 基质金属蛋白酶9 上游:CGGTCCTCACCATGAGTCC 162 下游:ACAAGTATGCCTCTGCCAGC 26型胶原纤维α1 上游:AGAGCGGTGCAAAATGAAGC 301 下游:GAGAGTCCTCACGAGGTTGG 无翅型MMTV整合位点家族成员6 上游:GATTTCTTCTCTCCAGCGAGC 252 下游:TGAGAGCTCTACCGCAGTCT 3-磷酸甘油醛脱氢酶 上游:GAGCGAGATCCCTCCAAAAT 197 下游:GCTGTTGTCATACTTCTCATGG  下载: 导出CSV
下载: 导出CSV
表2 实时荧光定量反转录PCR法检测毛乳头细胞生物功能标志物mRNA表达的引物序列及产物大小
引物名称 引物序列(5'→3') 产物大小(bp) 成纤维细胞生长因子7 上游:TATTCATGAACACCCGGGGC 168 下游:CAGTTCACACTCGTAGCCGT 无翅型MMTV整合位点家族成员10a 上游:TCAGACAGCTGGGGGTGATA 122 下游:TTGCCTATCACTCCTGGGGA 淋巴样增强因子1 上游:CTTCTCTGTCCCGATGGCAG 120 下游:TTGTTGTACAGGCCTCCGTC 碱性磷酸酶 上游:TCTGAAGGCTCTCTTCACTCC 75 下游:GCTACCAACTGCTGTGACCA β连环蛋白 上游:GGCGGCCGCGAGGTA 421 下游:TTAGTGGGATGAGCAGCGTC 性别决定区Y框蛋白2 上游:GGAGGAGAGCGCCTGTTTT 80 下游:CTGGCGGAGAATAGTTGGGG 多功能蛋白聚糖 上游:AGCTACGGAGCTCTTCAGGA 326 下游:AGATCACATAGGAAGCGCGG
下载: 导出CSV
-

 下载:
下载:




计量
- 文章访问数: 467
- HTML全文浏览量: 173
- PDF下载量: 26
- 被引次数: 0
