



Study on the effect and mechanism of recombinant human intestinal trefoil factor on intestinal mucosal injury and repair in burned mice
-
摘要:
目的 建立高效的人肠三叶因子(ITF)重组表达及纯化策略,观察重组人ITF(rhITF)对烧伤小鼠肠黏膜损伤与修复的作用并探讨其机制。 方法 采用实验研究方法。采用新型酵母表达载体pGAPZαA和酵母菌X33重组表达ITF,并通过金属螯合亲和层析与阴、阳离子交换层析法纯化蛋白,行非还原十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)及蛋白质印迹法鉴定rhITF。rhITF与胃蛋白酶溶液、胰蛋白酶溶液按体积比1∶1混合,分别作用0.5、1.0、1.5、2.0 h和1.0、2.0、4.0 h后行非还原SDS-PAGE,分析rhITF的稳定性。按随机数字表法将105只6~8周龄BALB/c雄性小鼠分为假伤组(30只)、单纯烧伤组(45只)、烧伤+rhITF组(30只)。将单纯烧伤组和烧伤+rhITF组小鼠背部造成30%体表总面积Ⅲ度烧伤,假伤组小鼠模拟致假伤,烧伤+rhITF组小鼠伤后灌胃1 mg/kg rhITF,其余2组小鼠伤后给予等量生理盐水。伤后24 h,取15只单纯烧伤组小鼠,制备烧伤血清,用于细胞实验。伤后3、5、7 d,每组各取10只小鼠处死后取小肠组织,苏木精-伊红染色观察肠黏膜病理变化,分光光度法和酶联免疫吸附测定法测定肠组织二胺氧化酶(DAO)及乳酸脱氢酶(LDH)活性。取3批人结直肠腺癌HT-29细胞,分为阴性对照组、25 μg/mL rhITF组、50 μg/mL rhITF组(样本数为3),正常对照组、烧伤血清组、烧伤血清+rhITF组(样本数为3),CK869抑制剂组、CK666抑制剂组和溶剂对照组(样本数为2),分别进行相应处理,Transwell实验观察培养12 h后细胞移行。另取2批HT-29细胞,每批细胞均分为正常对照组、烧伤血清组和烧伤血清+rhITF组(样本数均为6),培养24 h后,蛋白质印迹法检测细胞腺苷酸活化蛋白激酶(AMPK)、磷酸化AMPK(p-AMPK)、Ras相关C3肉毒杆菌毒素底物1(Rac1)及肌动蛋白相关蛋白2/3(Arp2/3)复合亚基1B(ARPC1B)蛋白表达,活化磁珠下拉实验检测细胞Rac1活性。对数据行析因设计方差分析、单因素方差分析、SNK检验。 结果 每升发酵液获得rhITF 82.35 mg,蛋白纯度高达98%,且具有良好的抗原特异性。rhITF在胃蛋白酶和胰蛋白酶溶液中较稳定,与胰蛋白酶溶液作用2.0 h后仍有45%残余,与胃蛋白酶溶液作用4.0 h后仍有90%残余。假伤组小鼠伤后各时间点肠黏膜未见充血、水肿,单纯烧伤组小鼠伤后各时间点肠黏膜主要病理表现为充血、水肿、糜烂及出血。烧伤+rhITF组小鼠伤后3、5 d肠黏膜主要以充血、水肿为主,伤后7 d肠黏膜水肿和充血减轻。与单纯烧伤组比较,假伤组、烧伤+rhITF组小鼠伤后3、5、7 d肠组织DAO和LDH活性显著升高(P<0.05或P<0.01)。培养12 h后,25 μg/mL rhITF组细胞移行数为(58±12)个,显著多于阴性对照组的(16±5)个(P<0.01),显著少于50 μg/mL rhITF组的(123±9)个(P<0.05)。培养12 h后,烧伤血清组细胞移行数为(60±13)个,显著少于正常对照组的(143±11)个和烧伤血清+rhITF组的(138±8)个(P<0.05)。培养12 h后,溶剂对照组细胞移行数为(155±9)个,明显多于CK666抑制剂组的(33±5)个、CK869抑制剂组的(28±5)个(P<0.01)。培养24 h后,烧伤血清组细胞AMPK、Rac1蛋白表达量与正常对照组、烧伤血清+rhITF组相近(P>0.05),p-AMPK蛋白表达量明显高于正常对照组、烧伤血清+rhITF组(P<0.05或P<0.01),ARPC1B蛋白表达量明显低于正常对照组、烧伤血清+rhITF组(P<0.05)。培养24 h后,烧伤血清组细胞Rac1活性明显低于正常对照组、烧伤血清+rhITF组(P<0.05或P<0.01)。 结论 本研究获得的rhITF具有较高的纯度和超强的稳定性,能耐受极端pH和蛋白酶水解,减轻烧伤小鼠肠黏膜损伤。rhITF能通过抑制AMPK磷酸化维持Rac1-Arp2/3活性,促进肠上皮细胞移行,加速肠黏膜修复。 Abstract:Objective To establish an efficient human intestinal trefoil factor (ITF) recombinant expression and purification strategy and to observe the effect of recombinant human ITF (rhITF) on intestinal mucosal injury and repair in burned rats and to explore the mechanism. Methods The experimental research method was applied. New yeast expression vector pGAPZαA and yeast X33 were used to express recombinant ITF. The protein was purified by metal chelation affinity chromatography and anion and cation exchange chromatography. The rhITF was identified by non-reductive sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and Western-blotting. The rhITF was mixed with pepsin solution and trypsin solution in a volume ratio of 1∶1, respectively. After mixed with pepsin solution for 0.5, 1.0, 1.5, 2.0 h and trypsin solution for 1.0, 2.0, 4.0 h, the stability of rhITF was analyzed with non-reductive SDS-PAGE. One hundred and five male BALB/c mice aged 6-8 weeks were divided into sham injury group (n=30), burn alone group (n=45), and burn+rhITF group (n=30) according to the random number table. Mice in burn alone group and burn+rhITF group were inflicted with 30% total body surface area full-thickness burn on the back, while mice in sham injury group were simulated with burn. After burn, mice in burn+rhITF group were intragastrically administered with rhITF of 1 mg/kg, while mice in the other two groups were given the same amount of normal saline. At post injury hour 24, 15 mice in burn alone group were collected to prepare burn serum, which was used in the cell experiment. On post injury day (PID) 3, 5, and 7, 10 mice in each group were sacrificed to collect the small intestinal tissue. The pathological changes of the intestinal mucosa were observed by hematoxylin-eosin staining, and the activities of diamine oxidase (DAO) and lactic dehydrogenase (LDH) in the intestinal tissue were determined by spectrophotometry and enzyme linked immunosorbent assay. Three batches of human colorectal adenocarcinoma HT-29 cells were taken and divided into negative control group, 25 μg/mL rhITF group, 50 μg/mL rhITF group (n=3), normal control group, burn serum group, burn serum+rhITF group (n=3), and CK869 inhibitor group, CK666 inhibitor group, solvent control group (n=2), respectively, which were dealt with the corresponding treatment. After 12 h of culture, the migration of cells were observed by Transwell experiment. Another 2 batches of HT-29 cells were taken and each batch of cells were divided into normal control group, burn serum group, and burn serum+rhITF group (n=6). After 24 h of culture, the protein expressions of adenosine monophosphate activated protein kinase (AMPK), phosphorylated AMPK (p-AMPK), Ras related C3 botulinum toxin substrate 1 (Rac1), and actin-related protein 2/3 (Arp 2/3) complex, subunit 1B (ARPC1B) in the cells were detected by Western blotting, and the Rac1 activity of the cells was detected by activated magnetic bead pull-down test. Data were statistically analyzed with analysis of variance for factorial design, one-way analysis of variance, and Student-Newman-Keuls test. Results Totally 82.35 mg rhITF was gathered from per litre of fermentation broth with protein purity up to 98%, and the rhITF had good antigenicity. The rhITF was stable in pepsin solution and trypsin solution, with 45% rhITF remained after 2.0 h in trypsin solution, and there was 90% rhITF remained after 4.0 h in pepsin solution. At each time point post injury, no hyperemia, or edema was observed in intestinal mucosa of mice in sham injury group, the main pathological manifestations of intestinal mucosa in mice of burn alone group were hyperemia, edema, erosion, and hemorrhage, and the main manifestations of intestinal mucosa of mice in burn+rhITF group were hyperemia and edema on PID 3 and 5, which were alleviated on PID 7. Compared with those of burn alone group, the activities of DAO and LDH in intestinal tissue of mice in sham injury group and burn+rhITF group were significantly increased on PID 3, 5, and 7 (P<0.05 or P<0.01 ). After 12 h of culture, the number of cell migration in 25 μg/mL rhITF group was 58±12, which was obviously more than 16±5 in negative control group (P<0.01) and obviously less than 123±9 in 50 μg/mL rhITF group (P<0.05). After 12 h of culture, the number of cell migration in burn serum group was 60±13, which was significantly less than 143±11 in normal control group and 138±8 in burn serum+rhITF group (P<0.05). After 12 h of culture, the number of cell migration in solvent control group was 155±9, which was significantly more than 33±5 in CK666 inhibitor group and 28±5 in CK869 inhibitor group (P<0.01). After 24 h of culture, the protein expressions of AMPK and Rac1 of cells in burn serum group were close to those of normal control group and burn serum+rhITF group (P˃0.05), the protein expression of p-AMPK of cells in burn serum group was significantly higher than that of normal control group and burn serum+rhITF group, respectively (P<0.05 or P<0.01), and the protein expression of ARPC1B of cells in burn serum group was significantly lower than that of normal control group and burn serum+rhITF group (P<0.05). After 24 h of culture, the Rac1 activity of cells in burn serum group was significantly lower than that in normal control group and burn serum+rhITF group, respectively (P<0.05 or P<0.01). Conclusions The rhITF obtained in this study has high purity and super stability, which can resist extreme pH and hydrolysis of protease and can relieve intestinal mucosal damage in burned mice. The rhITF can promote the migration of intestinal epithelial cells and accelerate the repair of intestinal mucosa through inhibiting phosphorylation of AMPK to maintain Rac1-Arp2/3 activity. -
Key words:
- Burns /
- Intestinal mucosa /
- Intestinal epithelial cells /
- Cell migration /
- Intestinal trefoil factor
-
参考文献
(20) [1] HeW,WangY,WangP,et al.Intestinal barrier dysfunction in severe burn injury[J/OL].Burns Trauma,2019,7:24[2021-04-12]. https://pubmed.ncbi.nlm.nih.gov/31372365/. DOI: 10.1186/s41038-019-0162-3. [2] QuirósM, NusratA. Contribution of wound-associated cells and mediators in orchestrating gastrointestinal mucosal wound repair[J]. Annu Rev Physiol, 2019, 81:189-209. DOI: 10.1146/annurev-physiol-020518-114504. [3] MatthewsJD, SumaginR, HinrichsB, et al. Redox control of Cas phosphorylation requires Abl kinase in regulation of intestinal epithelial cell spreading and migration[J]. Am J Physiol Gastrointest Liver Physiol, 2016,311(3):G458-465. DOI: 10.1152/ajpgi.00189.2016. [4] MoriyamaK,KouchiY,MorinagaH,et al.Diamine oxidase, a plasma biomarker in rats to GI tract toxicity of oral fluorouracil anti-cancer drugs[J].Toxicology,2006,217(2/3):233-239.DOI: 10.1016/j.tox.2005.09.017. [5] TakadaK,YamashitaK,Sakurai-YamashitaY,et al.Participation of nitric oxide in the mucosal injury of rat intestine induced by ischemia-reperfusion[J].J Pharmacol Exp Ther,1998,287(1):403-407. [6] WangZE,PengJ,WuD,et al.Effects of intestinal trefoil factor on intestinal mucus barrier in burned mice[J].Am J Transl Res,2020,12(11):7187-7198. [7] MashimoH,WuDC,PodolskyDK,et al.Impaired defense of intestinal mucosa in mice lacking intestinal trefoil factor[J].Science,1996,274(5285):262-265.DOI: 10.1126/science.274.5285.262. [8] HoffmannW.Trefoil factor family (TFF) peptides: regulators of mucosal regeneration and repair, and more[J].Peptides,2004,25(5):727-730.DOI: 10.1016/j.peptides.2004.03.019. [9] SunY,PengX,ZhangY,et al.Stability and biological activity of human intestinal trefoil factor produced by Pichia pastoris[J].Protein Pept Lett,2008,15(3):255-259.DOI: 10.2174/092986608783744243. [10] LeJ,ZhangDY,ZhaoY,et al.ITF promotes migration of intestinal epithelial cells through crosstalk between the ERK and JAK/STAT3 pathways[J].Sci Rep,2016,6:33014.DOI: 10.1038/srep33014. [11] KlocM,GhobrialRM.Chronic allograft rejection: a significant hurdle to transplant success[J/OL].Burns Trauma,2014,2(1):3-10[2021-04-12]. https://pubmed.ncbi.nlm.nih.gov/27574640/. DOI: 10.4103/2321-3868.121646. [12] KastDJ, DominguezR. IRSp53 coordinates AMPK and 14-3-3 signaling to regulate filopodia dynamics and directed cell migration[J]. Mol Biol Cell, 2019, 30(11):1285-1297. DOI: 10.1091/mbc.E18-09-0600. [13] KrndijaD,El MarjouF,GuiraoB,et al.Active cell migration is critical for steady-state epithelial turnover in the gut[J].Science,2019,365(6454):705-710.DOI: 10.1126/science.aau3429. [14] SwaneyKF,LiR.Function and regulation of the Arp2/3 complex during cell migration in diverse environments[J].Curr Opin Cell Biol,2016,42:63-72.DOI: 10.1016/j.ceb.2016.04.005. [15] O'NeilSD, RáczB, BrownWE, et al. Action potential-coupled Rho GTPase signaling drives presynaptic plasticity[J]. Elife, 2021,10:e63756. DOI: 10.7554/eLife.63756. [16] KunitomiH, OkiY, OnishiN, et al. The insulin-PI3K-Rac1 axis contributes to terminal adipocyte differentiation through regulation of actin cytoskeleton dynamics[J]. Genes Cells, 2020,25(3):165-174. DOI: 10.1111/gtc.12747. [17] NakanoA, KatoH, WatanabeT, et al. AMPK controls the speed of microtubule polymerization and directional cell migration through CLIP-170 phosphorylation[J]. Nat Cell Biol, 2010 ,12(6):583-590. DOI: 10.1038/ncb2060. [18] BaeHB, ZmijewskiJW, DeshaneJS, et al. AMP-activated protein kinase enhances the phagocytic ability of macrophages and neutrophils[J]. FASEB J, 2011,25(12):4358-4368. DOI: 10.1096/fj.11-190587. [19] LeeYM, LeeJO, JungJH,et al. Retinoic acid leads to cytoskeletal rearrangement through AMPK-Rac1 and stimulates glucose uptake through AMPK-p38 MAPK in skeletal muscle cells[J]. J Biol Chem, 2008, 283(49):33969-33974. DOI: 10.1074/jbc.M804469200. [20] YanY, TsukamotoO, NakanoA, et al. Augmented AMPK activity inhibits cell migration by phosphorylating the novel substrate Pdlim5[J]. Nat Commun, 2015,6:6137. DOI: 10.1038/ncomms7137. -

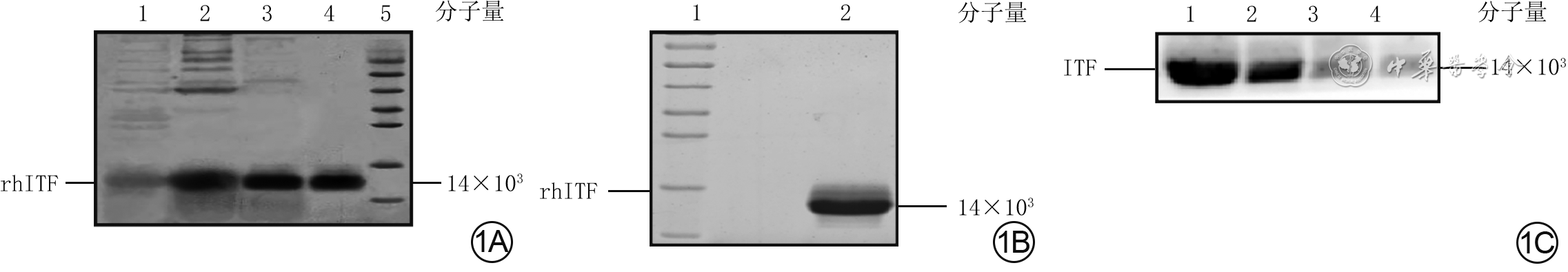
1 非还原十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)检测人ITF重组表达及蛋白质印迹法检测其抗原特异性。1A.层析样品非还原SDS-PAGE图谱,1为rhITF发酵上清液,2为阳离子交换层析后的蛋白溶液,3为金属螯合亲和层析后的蛋白溶液,4为阴离子交换层析后的蛋白溶液,5为蛋白质预染marker;1B.经3个步骤纯化后的rhITF SDS-PAGE图谱,1为蛋白质预染marker;2为rhITF;1C.蛋白质印迹法检测rhITF的蛋白表达,1为40 μg重组蛋白,2为20 μg重组蛋白,3为10 μg重组蛋白,4为5 μg重组蛋白
注:ITF为肠三叶因子,rhITF为重组人ITF

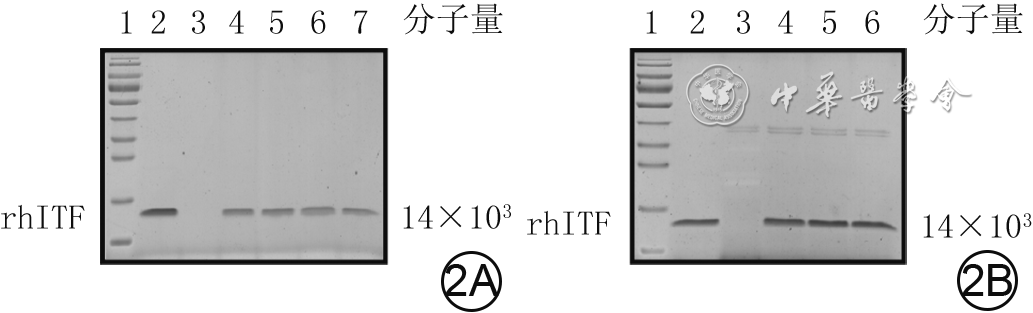
2 非还原十二烷基硫酸钠-聚丙烯酰胺凝胶电泳检测rhITF在胰蛋白酶和胃蛋白酶溶液中的稳定性。2A.rhITF在胰蛋白酶溶液中的稳定性,1为蛋白质预染marker,2为rhITF,3为500 mg/L胰蛋白酶溶液,4~7分别为rhITF与500 mg/L胰蛋白酶溶液作用0.5、1.0、1.5、2.0 h;2B.rhITF在胃蛋白酶溶液中的稳定性,1为蛋白质预染marker,2为rhITF,3为500 mg/L胃蛋白酶溶液,4~6分别为rhITF与500 mg/L胃蛋白酶溶液作用1.0、2.0、4.0 h
注:rhITF为重组人肠三叶因子

3 假伤组及烧伤2组小鼠伤后3、5 d小肠黏膜病理变化 苏木精-伊红×400,图中标尺为50 μm。3A.假伤组伤后3 d肠黏膜未见充血、水肿;3B、3C.分别为单纯烧伤组、烧伤+重组人肠三叶因子(rhITF)组伤后3 d小肠病理变化,单纯烧伤组可见肠黏膜充血、水肿,肠上皮细胞空泡变性、肠绒毛萎缩,烧伤+rhITF组肠黏膜可见充血、水肿;3D.假伤组伤后5 d肠黏膜病理变化与图3A相近;3E、3F.分别为单纯烧伤组、烧伤+rhITF组伤后5 d肠黏膜病理变化,单纯烧伤组仍可见肠黏膜充血、水肿,烧伤+rhITF组肠黏膜充血、水肿较图3C减轻
注:黑色箭头示肠上皮细胞空泡变性、肠绒毛萎缩,白色箭头示肠黏膜充血、水肿

4 阴性对照组及重组人肠三叶因子(rhITF)处理2组HT-29细胞培养12 h后移行情况 结晶紫×200,图中标尺为100 μm。4A.阴性对照组细胞移行数较少;4B.25 μg/mL rhITF组细胞移行数多于图4A;4C.50 μg/mL rhITF组细胞移行数多于图4A、4B

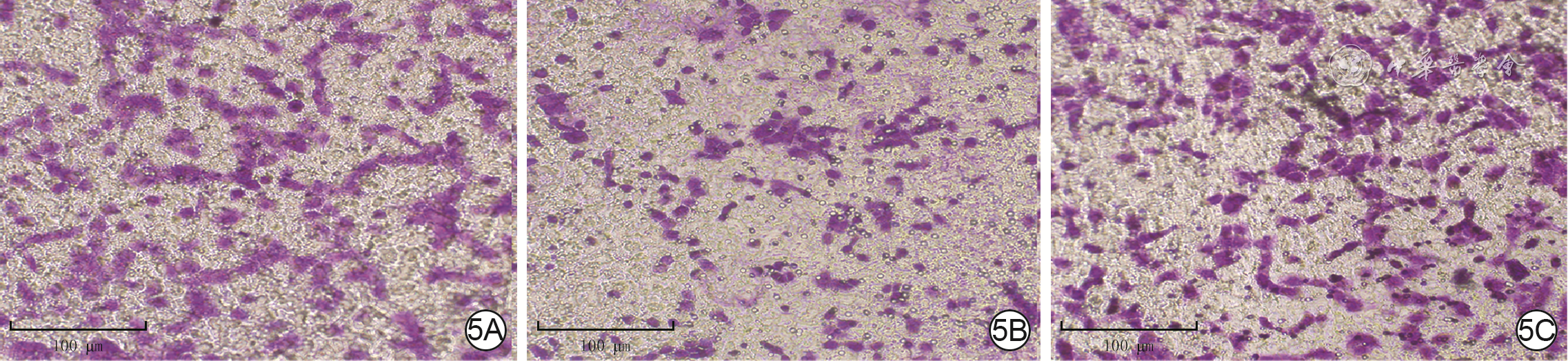
5 3组HT-29细胞培养12 h后移行情况 结晶紫×200,图中标尺为100 μm。5A.正常对照组细胞移行数较多;5B.烧伤血清组细胞移行数少于图5A;5C.烧伤血清+重组人肠三叶因子组细胞移行数多于图5B

6 溶剂对照组及肌动蛋白相关蛋白2/3复合物抑制剂处理2组HT-29细胞培养12 h后移行情况 结晶紫×200,图中标尺为100 μm。6A.溶剂对照组移行细胞较多;6B、6C.分别为CK666抑制剂组和CK869抑制剂组,2组移行细胞数相近,且均较图6A减少

7 蛋白质印迹法检测3组HT-29细胞培养24 h后的Rac1-Arp2/3信号通路相关蛋白表达及活化磁珠下拉检测Rac1活性。7A.Rac1、p-AMPK、AMPK、ARPC1B蛋白表达;7B.GTP-Rac1蛋白表达反映Rac1活性
注:1.正常对照组,2.烧伤血清组,3.烧伤血清+重组人肠三叶因子组;Rac1为Ras相关C3肉毒杆菌毒素底物1,Arp2/3为肌动蛋白相关蛋白2/3,AMPK为腺苷酸活化蛋白激酶,p-AMPK为磷酸化AMPK,ARPC1B为Arp2/3复合亚基1B,GAPDH为3-磷酸甘油醛脱氢酶,GTP-Rac1为Rac1与三磷酸鸟苷复合物
表1 假伤组及烧伤2组小鼠伤后各时间点小肠组织二胺氧化酶及乳酸脱氢酶活性比较(U/mg,
组别与时间点 鼠数(只) 二胺氧化酶 乳酸脱氢酶 假伤组 30 伤后3 d 60.7±3.4 55.1±1.4 伤后5 d 63.5±1.4 75.7±3.8 伤后7 d 67.5±5.3 62.5±4.2 单纯烧伤组 30 伤后3 d 33.7±4.9a 10.4±1.6a 伤后5 d 34.9±4.1a 16.3±2.9a 伤后7 d 43.6±3.6a 14.9±2.6a 烧伤+重组人肠三叶因子组 30 伤后3 d 48.6±2.4b 34.2±4.1b 伤后5 d 51.2±3.3b 49.0±2.4b 伤后7 d 55.3±4.0b 47.2±3.2b F1值 95.39 131.80 P1值 <0.01 <0.01 F2值 186.90 264.50 P2值 <0.01 <0.01 F3值 22.79 70.79 P3值 <0.01 <0.01 注:各组各时间点鼠数为10只;二胺氧化酶活性及乳酸脱氢酶活性处理因素主效应,F=256.31、92.45,P<0.01;时间因素主效应,F=88.27、169.80,P<0.01;两者交互作用,F=23.64、45.28,P<0.01;F1值、P1值,F2值、P2值,F3值、P3值分别为3组伤后3、5、7 d各指标总体比较所得;与假伤组比较,aP<0.01;与单纯烧伤组比较,bP<0.05  下载: 导出CSV
下载: 导出CSV
表2 3组HT-29细胞培养24 h Rac1-Arp2/3信号通路相关蛋白表达比较(
组别 样本数 AMPK p-AMPK ARPC1B Rac1 GTP-Rac1 正常对照组 6 1.000±0.032 1.00±0.03 1.00±0.04 1.00±0.11 1.000±0.054 烧伤血清组 6 0.962±0.023 2.04±0.07a 0.56±0.03c 1.17±0.04 0.564±0.041a 烧伤血清+重组人肠三叶因子组 6 1.043±0.021 1.51±0.08b 0.81±0.03b 1.12±0.03 0.831±0.023b F值 9.45 175.21 96.98 4.69 93.50 P值 >0.05 <0.01 <0.01 >0.05 <0.01 注:Rac1为Ras相关C3肉毒杆菌毒素底物1,Arp2/3为肌动蛋白相关蛋白2/3,AMPK为腺苷酸活化蛋白激酶,p-AMPK为磷酸化AMPK,ARPC1B为Arp2/3复合亚基1B,GTP-Rac1为Rac1与三磷酸鸟苷复合物、F值、P值为3组间各指标总体比较所得;与正常对照组比较,aP<0.01,cP<0.05;与烧伤血清组比较,bP<0.05
下载: 导出CSV
-

 下载:
下载:





计量
- 文章访问数: 367
- HTML全文浏览量: 151
- PDF下载量: 22
- 被引次数: 0
