



The effect and mechanism of exosomes derived from human amniotic epithelial cells on the proliferation and migration of HaCaT in high glucose environment
-
摘要:
目的 探讨人羊膜上皮细胞外泌体(hAEC-Exo)对高糖环境下HaCaT增殖和迁移的作用及其相关机制。 方法 采用实验研究方法。取2019年1—6月于福建医科大学附属协和医院妇产科10名足月分娩健康孕妇羊膜组织,分离原代人羊膜上皮细胞(hAEC)。观察培养第2、4、7天原代hAEC生长状态和形态改变,采用流式细胞术检测细胞表面标志物CD73、CD90、CD29、CD34及人白细胞抗原DR(HLA-DR)的表达,取第2~4代细胞用于后续实验。超速离心法分离hAEC-Exo。将HaCaT与hAEC-Exo共培养3 h,采用倒置荧光显微镜观察HaCaT对hAEC-Exo的摄取情况。取HaCaT,分为磷酸盐缓冲液(PBS)组、hAEC-Exo组和二甲基亚砜(DMSO)+PBS组、DMSO+hAEC-Exo组、LY294002+hAEC-Exo组,每组3孔,并进行相应处理,采用细胞计数试剂盒8(CCK-8)法检测培养0(即刻)、12、24、36、48、60 h细胞增殖活力;划痕试验检测划痕后0、24、48、72 h划痕愈合情况,并计算划痕愈合率;Transwell实验检测培养48 h穿膜细胞数;蛋白质印迹法检测培养24 h后磷脂酰肌醇3-激酶-蛋白激酶B-哺乳动物雷帕霉素靶蛋白(PI3K-Akt-mTOR)通路相关的哺乳动物雷帕霉素靶蛋白(mTOR)、磷酸化mTOR(p-mTOR)、蛋白激酶B(Akt)、磷酸化Akt(p-Akt)的蛋白表达。对数据行重复测量方差分析、单因素方差分析及独立样本t检验。 结果 原代hAEC培养第2天多数呈卵圆形,大小均一;培养第4、7天,细胞形态呈典型的鹅卵石样单层排列。原代hAEC高表达间充质干细胞表面标志物CD73、CD90及CD29,不表达或低表达造血干细胞相关表面标志物CD34及HLA-DR。培养3 h,hAEC-Exo成功被HaCaT内吞入细胞质,并聚集于细胞核周围。培养12、24、36、48、60 h,hAEC-Exo组HaCaT增殖活力明显高于PBS组(t=3.691、10.861、12.121、10.531、14.931,P<0.01)。划痕后24、48、72 h,PBS组HaCaT划痕愈合率明显低于hAEC-Exo组(t=3.342、6.427、5.485,P<0.05或P<0.01)。培养48 h,hAEC-Exo组HaCaT穿膜数显著多于PBS组(t=5.385,P<0.01)。培养24 h,hAEC-Exo组HaCaT中p-mTOR和p-Akt蛋白表达量明显高于PBS组(t=4.240、5.586,P<0.01),2组HaCaT中mTOR和Akt蛋白表达量相近(P>0.05)。培养24 h,DMSO+hAEC-Exo组HaCaT中p-mTOR和p-Akt的蛋白表达量明显高于DMSO+PBS组(t=6.155、8.338,P<0.01)和LY294002+hAEC-Exo组(t=5.030、3.960,P<0.01),3组HaCaT中mTOR和Akt蛋白表达量相近(P>0.05)。DMSO+hAEC-Exo组HaCaT培养12、24、36、48、60 h增殖活力为0.78±0.05、1.23±0.07、1.60±0.09、1.86±0.09、2.03±0.08,明显高于DMSO+PBS组的0.46±0.04、0.69±0.07、0.98±0.08、1.16±0.08、1.26±0.11(t=4.376、7.398、8.488、9.766、10.730,P<0.01);DMSO+hAEC-Exo组HaCaT培养24、36、48、60 h增殖活力明显高于LY294002+hAEC-Exo组的0.96±0.09、1.20±0.08、1.39±0.08、1.55±0.10(t=3.639、5.447、6.605、6.693,P<0.05或P<0.01)。DMSO+hAEC-Exo组HaCaT划痕后24、48、72 h划痕愈合率明显高于DMSO+PBS组(t=4.003、6.349、7.714,P<0.01)和LY294002+hAEC-Exo组(t=3.805、4.676、4.067,P<0.05或P<0.01)。培养48 h,DMSO+hAEC-Exo组HaCaT穿膜数明显多于DMSO+PBS组和LY294002+hAEC-Exo组(t=7.464、1.232,P<0.01)。 结论 PI3K-Akt-mTOR通路介导hAEC-Exo促进高糖环境下HaCaT增殖和迁移。 Abstract:Objective To investigate the effect and mechanism of exosomes derived from human amniotic epithelial cells (hAEC-Exos) on the proliferation and migration of HaCaT in high glucose environment. Methods The experimental research method was adopted. The amniotic membrane tissue was collected from 10 healthy pregnant women at full term delivery in the Department of Obstetrics and Gynecology of Fujian Medical University Union Hospital from January to June 2019, and the primary human amniotic epithelial cells (hAECs) were isolated. The growth status and morphological changes of the primary hAECs on the 2nd, 4th, and 7th day of culture were observed, and the expressions of the cells surface markers of CD73, CD90, CD29, CD34, and human leukocyte antigen DR (HLA-DR). The 2nd to 4th passages of hAECs were used in the following experiments. The hAEC-Exos were separated by ultracentrifugation method. The HaCaT and hAEC-Exos were co-cultured for 3 h, and the uptake of hAEC-Exos by HaCaT was observed by inverted fluorescence microscopy. The HaCaT were divided into phosphate buffer solution (PBS) group and hAEC-Exos group or dimethyl sulfoxide (DMSO)+PBS group, DMSO+hAEC-Exos group, and LY294002+hAEC-Exos group, which were dealt correspondingly, with 3 wells in each group. Cell counting kit 8 (CCK-8) method was used to detect cell proliferation activity after 0 (immediately), 12, 24, 36, 48, and 60 h of culture. The scratch test was conducted to detect the scratch healing at 0, 24, 48, and 72 h after the scratch, and the scratch healing rate was calculated, respectively. The Transwell experiment was conducted to detect the number of transmembrane cells after 48 h of culture. The Western blotting was used to detect the protein expressions of mammalian target of rapamycin (mTOR), phosphorylated mTOR (p-mTOR), protein kinase B (Akt), and phosphorylated Akt (p-Akt) related to phosphatidylinositol 3-kinase-Akt-mTOR (PI3K-Akt-mTOR) pathway after 24 h of culture. Data were statistically analyzed with analysis of variance for repeated measurement, one-way analysis of variance, and independent sample t test. Results Most of the primary hAECs were oval and uniform in size on the 2nd day of culture. The hAECs were arranged in a typical cobblestone-like monolayer on the 4th and 7th day of culture. The primary hAECs highly expressed CD73, CD90, and CD29 of mesenchymal stem cell related surface markers, and were with no or low expressions of CD34 and HLA-DR of hematopoietic stem cell related surface markers. After 3 h of culture, hAEC-Exos were successfully endocytosed by HaCaT into the cytoplasm and gathered around the nucleus. After 12, 24, 36, 48, and 60 h of culture, the proliferation activity of HaCaT in hAEC-Exos group was significantly higher than that in PBS group (t=3.691, 10.861, 12.121, 10.531, 14.931, P<0.01). At 24, 48, and 72 h after scratch, the scratch healing rates of HaCaT in PBS group were significantly lower than those in hAEC-Exos group (t=3.342, 6.427, 5.485, P<0.05 or P<0.01). After 48 h of culture, the number of transmembrane HaCaT in hAEC-Exos group was significantly more than that in PBS group (t=5.385, P<0.01). After 24 h of culture, the protein expressions of p-mTOR and p-Akt in HaCaT of hAEC-Exos group were significantly higher than those in PBS group (t=4.240, 5.586, P<0.01), while the protein expressions of mTOR and Akt in HaCaT of the two groups were similar (P>0.05). After 24 h of culture, the protein expressions of p-mTOR and p-Akt in HaCaT of DMSO+hAEC-Exos group were significantly higher than those in DMSO+PBS group (t=6.155, 8.338, P<0.01) and LY294002+hAEC-Exos group (t=5.030, 3.960, P<0.01), while the protein expressions of mTOR and Akt in HaCaT of the three groups were similar (P>0.05). The proliferation activity of HaCaT in DMSO+hAEC-Exos group at 12, 24, 36, 48, and 60 h of culture was 0.78±0.05, 1.23±0.07, 1.60±0.09, 1.86±0.09, and 2.03±0.08, which was significantly higher than 0.46±0.04, 0.69±0.07, 0.98±0.08, 1.16±0.08, and 1.26±0.11 in DMSO+PBS group (t=4.376, 7.398, 8.488, 9.766, 10.730, P<0.01). The proliferation activity of HaCaT in DMSO+hAEC-Exos group at 24, 36, 48, and 60 h was significantly higher than 0.96±0.09, 1.20±0.08, 1.39±0.08, and 1.55±0.10 in LY294002+hAEC-Exos group (t=3.639, 5.447, 6.605, 6.693, P<0.05 or P<0.01). The scratch healing rates of HaCaT in DMSO+hAEC-Exos group at 24, 48, and 72 h after scratch were significantly higher than those in DMSO+PBS group (t=4.003, 6.349, 7.714, P<0.01) and LY294002+hAEC-Exos group (t=3.805, 4.676, 4.067, P<0.05 or P<0.01). After 48 h of culture, the number of transmembrane HaCaT in DMSO+hAEC-Exos group was significantly more than that in DMSO+PBS group and LY294002+hAEC-Exos group, respectively (t=7.464, 1.232, P<0.01). Conclusions PI3K-Akt-mTOR pathway can promote the proliferation and migration of HaCaT in high glucose environment by mediating hAEC-Exos. -
烧创伤导致的严重体表组织缺损对患者打击巨大,除容易引发感染和脓毒症外,还会导致骨、肌肉等深部组织外露甚至进行性坏死,形成更复杂的难治性创面。快速发展的急救技术显著提高了烧创伤患者的抢救成功率,但烧创伤创面的治疗仍存在难题。首先是创面瘢痕愈合,虽然生命得以挽救,患者仍长期遭受瘢痕带来的困扰,包括毁容、挛缩、瘙痒、无法排汗等。烧伤创面患者增生性瘢痕发生率高达70%[1]。据估计,全球范围每年用于瘢痕治疗的医疗费用在2027年将达到320亿美元,是沉重的社会负担[2]。此外,生存质量的下降还给患者造成沉重的心理负担[3]。其次是复杂创面的修复难题。由于创基条件差,胸骨、肌肉等深部组织外露创面的治疗十分棘手。在感染等因素影响下这些创面会转变为慢性创面,进一步加大治疗难度,最终会导致截肢等。解决这些难题具有重要的临床价值,也一直是创面修复领域的研究热点。结合本课题组的研究经验,本文主要总结了近期国内外有关生物活性材料促进创面修复和皮肤附件再生的研究成果和进展,为后续研究提供参考。
1. 创面微环境与生物活性材料的关系
创面愈合是人体在生物进化过程中获得的一种自我保护功能,使人体免遭外界污染物的危害而得以存活。但是,创面自愈合能力有限,自然状态下,烧创伤创面愈合缓慢,增加进行性加深和感染的风险。而且,自愈创面与临床干预创面相比更容易发生异常愈合。创面深度过深、张力过高、异常炎症反应等均是引发异常愈合,尤其是瘢痕愈合的原因。与成人相比,人类妊娠早期胎儿创面发生无瘢痕愈合。成人和早期胎儿创面微环境差异被认为是决定创面不同愈合模式的关键因素[4]。
创面微环境是指创面局部及其邻近区域中影响创面愈合的物理、生物化学、细胞等因素的总和,可分为外部微环境和内部微环境[5]。外部微环境涵盖温度、湿度、张力、酸碱度等因素,内部微环境涵盖ECM和细胞因素。不同原因影响创面愈合的作用最终均体现于对内部微环境的改变,例如,创面张力通过提高ECM刚度促进肌Fb(MFb)分化[6]。不利于创面再生性修复的内部微环境因素包括异常的炎症反应、生物化学和生物物理信号、细胞表型和功能。调控创面微环境的本质是通过材料、药物、物理疗法等不同方法直接或间接地改善内部微环境,在生物化学、机械力、空间结构等多方面调控免疫和非免疫细胞行为,进而影响创面愈合过程。微环境调控是促进创面再生修复的必由之路[7]。
生物活性材料在调控创面微环境方面具有显著优势。与局部用药、负压吸引等方法相比,利用生物活性材料可同时实现生物化学、机械力、空间结构等微环境要素的调节和营造,为创面细胞提供三维生长空间的同时,向细胞传递机械力、生物化学等信号。随着第3代生物材料理论体系的建立,创面修复生物活性材料的研发思路发生改变,通过再生医学、材料学、工程学等学科的交叉融合,构建主动、高效调控创面细胞行为的生物活性材料成为研究焦点[8]。近年来,以创面再生性修复和皮肤附件再生为目标,国内外研究者研发了系列新型生物活性材料,为创面修复提供新材料的同时,进一步加深了人们对创面修复的理解。
2. 生物活性材料促进创面修复和皮肤再生的机制
2.1 影响免疫细胞行为
利用生物活性材料可实现对创面免疫细胞行为和炎症反应的调控。损伤修复和组织重建的过程与炎症反应密切相关,因为免疫细胞不仅发挥吞噬病原体和坏死组织的作用[9],还通过分泌细胞因子、生长因子以及基质金属蛋白酶(MMP)等参与组织重建过程。一些生物活性材料通过诱导巨噬细胞发生M2型极化,营造促再生炎症反应微环境。葡聚糖-甲基丙烯酸异氰基乙酯-乙胺水凝胶诱导巨噬细胞发生M2型极化,其促进真皮功能性重建和毛囊再生的作用分别在小鼠和猪全层皮肤缺损创面模型中得到验证[10]。膀胱脱细胞基质(urinary bladder-decellularized matrix,UBM)和小肠黏膜下层基质等猪组织脱细胞基质诱导巨噬细胞向M2型极化,在烧创伤急性创面、下肢复杂性创面的修复中显示出与胶原蛋白基合成真皮替代物相比更强的促愈合能力[11, 12]。笔者课题组研究显示,糖尿病小鼠全层皮肤缺损创面巨噬细胞较快迁入并广泛分布于UBM中,UBM诱导迁入的巨噬细胞发生M2型极化,表达FGF-2、VEGF等促愈和促血管新生生长因子[13]。脱细胞基质材料含有基质结合纳米囊,后者含有的微小RNA被证实是调控巨噬细胞向抗炎表型极化的关键分子[14]。临床证据与上述结论相符,UBM单独应用即可促进肌腱和骨外露复杂创面愈合和真皮重建[15],而胶原蛋白基合成真皮替代物需联合VSD系统以提高复杂创面愈合速度[16]。虽然缺少临床组织学证据,但大量的体外和动物模型研究证据提示,调控巨噬细胞行为并产生促再生微环境是ECM材料增强创面愈合的关键机制之一。
除调控炎症反应之外,近期研究证实了生物活性材料诱导创面发生适应性免疫反应的功能。Jennifer Elisseeff课题组较早注意到UBM激活适应性免疫反应的作用[17],并证实UBM诱导小鼠角膜上皮缺损创面发生适应性免疫反应,促进角膜再生和视力恢复[18]。此外,一些合成生物活性材料也被证实具有激活创面适应性免疫反应的功能。Griffin等[19]对比了分别含有D型和L型氨基酸多肽的微孔退火颗粒水凝胶对小鼠全层皮肤缺损创面炎症反应的影响。结果显示,与L型相比,含有D型氨基酸多肽的微孔退火颗粒水凝胶可显著增强IL-33+骨髓细胞的募集,抑制瘢痕形成,并促进毛囊、皮脂腺再生。目前,与炎症反应相比,生物活性材料调控适应性免疫反应的机制有待明确,主要包括抗原产生、呈递过程及其与材料组成、结构等性质的关系[20]。
2.2 调控非免疫细胞行为
利用生物活性材料可实现对创面非免疫细胞行为的调控。在高张力、真皮缺失等因素的影响下,深度创面中的MFb过度活跃,大量合成胶原蛋白等基质分子,最终形成瘢痕组织。因此,调控创基细胞行为,抑制其向MFb分化是减轻瘢痕形成的重要策略[21]。然而,单独应用药物或物理的方法难以达到这一目的,因为调控MFb行为的因素多且复杂,不仅涉及炎症反应,还涉及细胞所处微环境的结构、力学性能和生物化学组成等。生物活性材料在调控MFb行为方面具有显著优势。已上市的Integra®、PELNAC®等胶原蛋白基真皮替代物显示出抑制瘢痕形成的功能,其关键作用机制之一即是通过支架中GFOGER和GLOGER多肽调控MFb的黏附行为,抑制MFb聚集及其收缩创面的作用[22]。但是,Integra®等真皮替代物未显示出对MFb分化的抑制性,而一些脱细胞基质材料则显示出阻碍创面MFb分化的作用[18]。例如,UBM抑制小鼠角膜上皮缺损创面MFb分化[18]。人羊膜脱细胞基质抑制兔耳全层皮肤缺损创面MFb分化,可降低胶原蛋白表达量和沉积量并提高MMP-1的表达和分泌,有效促进皮肤再生[23]。利用载药生物活性材料调控创面细胞行为是促进创面再生性修复的另一途径。含高渗葡萄糖酸钾的胶原蛋白水凝胶可抑制小鼠全层皮肤缺损创面MFb分化,促进皮肤再生[24]。载氧化锌-姜黄素纳米复合物的胶原蛋白支架可显著上调大鼠烧伤创面细胞表达的TGF-β3的活性,促进创面无瘢痕愈合[25]。含TGF-β1小干扰RNA的胶原蛋白-壳聚糖支架可长效抑制猪全层皮肤缺损创面细胞TGF-β1信号通路,促进皮肤再生[26]。
利用生物活性材料的空间结构和力学性能可实现对细胞存活、分化、空间排布等行为的调控。聚己内酯电纺丝支架与聚己内酯多孔支架相比,显示出促进脂肪间充质干细胞表达核因子κB的作用,增强细胞旁分泌功能,使大鼠全层皮肤缺损创面中M2型巨噬细胞增加[27]。笔者课题组构建了柔性丝蛋白纳米纤维支架,研究显示仅通过纳米结构和机械特性的优化即可诱导大鼠骨髓间充质干细胞向内皮细胞分化[28]。Xu等[29]利用聚乳酸和生物玻璃构成的纳米纤维支架向共培养的Fb和内皮细胞同时传递结构信号和生物化学信号,显著增强2种细胞的旁分泌功能,促进小鼠全层皮肤缺损创面血管化和再生性修复。生物活性材料通过黏附受体整合素、机械门控离子通道Piezo1蛋白等受体向细胞施加力学和空间结构的调控作用。在材料应力刺激下,巨噬细胞、Fb的整合素受体被激活,使潜在TGF-β1得到释放[30]。另外,生物活性材料通过肌动蛋白微丝连接整合素的瞬时受体电位M7和C1在细胞膜张力增加时促进钙离子内流,将力学刺激转化为电化学信号,影响细胞行为[31]。对接种于聚丙烯酰胺水凝胶的小鼠骨髓来源巨噬细胞,细胞Piezo1受体与整合素相比在应力作用下直接向胞内输送钙离子[32]。材料空间结构对细胞行为的影响是基于细胞黏附配体的空间分布和密度而实现的。在材料刚度一定的条件下,特定微结构如纳米纤维结构、取向结构、孔结构可改变力学刺激的方向和强度,进而调控细胞迁移、增殖和分化等行为[33];细胞间连接及其相互作用也受到材料空间结构的影响[33]。
2.3 促进皮肤附件再生
此外,生物活性材料在皮肤附件再生医学技术中显示出很高的应用潜能。首先,特定小分子药物联合生物可降解三维支架通过多因素协同作用促进皮肤附件再生。Geoffrey Gurtner团队以普鲁兰多糖-胶原蛋白复合水凝胶作为局部黏着斑激酶(focal adhesion kinase,FAK)抑制剂(化合物VS6062)的载体,显著增强杜洛克猪深度创面愈合作用的同时,实现了毛囊和汗腺的再生[34]。抑制FAK使Fb蛋白激酶B(Akt)信号放大,进而上调早期生长应答因子1 (EGR1)和乳脂球EGF8(MFGE8)这2种蛋白的表达,Akt-EGR1信号通路被认为与细胞再生性表型有关,而MFGE8具有促进胶原蛋白分子被吞噬的作用[35]。其次,通过化学手性异构激活适应性免疫反应的D型氨基酸多肽的微孔退火颗粒水凝胶支架可诱导小鼠全层皮肤缺损创面毛囊新生,此外,研究者进一步证实了毛囊新生对支架激活适应性免疫的依赖性[19],但相关分子机制有待明确。
生物活性材料与三维生物打印技术的结合为皮肤附件再生提供了新方案。以明胶和透明质酸衍生化合物作为三维生物打印墨水,可维持人Fb和人脐静脉内皮细胞团块在支架中的空间排布,维持打印组织的活力,促进大鼠和猪全层皮肤缺损创面血管化、再生性修复和毛囊、皮脂腺等附件再生[36]。以脱细胞基质来源材料作为生物墨水成分,可显著增强细胞对细胞因子、生长因子信号的响应性,为皮肤附件再生提供基础[37]。2020年,付小兵院士团队将足趾垫匀浆蛋白和生物墨水共同作为骨髓间充质干细胞的载体,应用三维打印技术在小鼠足趾垫烫伤创面实现了汗腺的原位再生,并证实了足趾匀浆蛋白在三维环境下诱导骨髓间充质干细胞表达胶原三螺旋重复蛋白1和血红素加氧酶基因1,促进汗腺再生[38]。
3. 总结与展望
综上,生物活性材料已发展成为创面修复技术的基础性要素[39]。近年来,系列主动调控创面微环境与细胞的新型生物活性材料得以研发,其调控创面愈合的新机制进一步加深了人们对创面修复的理解,为其临床应用提供了坚实的理论依据。然而,至今,大部分生物活性材料都是针对创面愈合的单一微环境要素或时间而设计和应用的。为了最终达到创面再生性修复的目的,生物活性材料应朝着整合多种作用机制的多功能材料发展,更有效地发挥其通过不同特性协同调控创面愈合的优势。因此,如何在同一体系中整合调控炎症反应、呈递适宜生物化学和生物物理信号的材料基元是关键的科学问题。此外,生物活性材料的研发和应用应更加重视创面的精准修复,针对创面的不同特点、调控创面微环境的不同目的和时机优化材料性能。相信随着再生医学、生物材料学和组织工程学的快速发展,将产生更多的新型生物活性材料[40],推动创面修复技术朝着精准、高效调控创面微环境的方向发展,为实现创面“完美修复”和皮肤再生奠定基础。
脱细胞真皮基质(ADM) 重症监护病房(ICU) 动脉血氧分压(PaO2) 丙氨酸转氨酶(ALT) 白细胞介素(IL) 磷酸盐缓冲液(PBS) 急性呼吸窘迫综合征(ARDS) 角质形成细胞(KC) 反转录-聚合酶链反应(RT-PCR) 天冬氨酸转氨酶(AST) 半数致死烧伤面积(LA50) 全身炎症反应综合征(SIRS) 集落形成单位(CFU) 内毒素/脂多糖(LPS) 超氧化物歧化酶(SOD) 细胞外基质(ECM) 丝裂原活化蛋白激酶(MAPK) 动脉血氧饱和度(SaO2) 表皮生长因子(EGF) 最低抑菌浓度(MIC) 体表总面积(TBSA) 酶联免疫吸附测定(ELISA) 多器官功能障碍综合征(MODS) 转化生长因子(TGF) 成纤维细胞(Fb) 多器官功能衰竭(MOF) 辅助性T淋巴细胞(Th) 成纤维细胞生长因子(FGF) 一氧化氮合酶(NOS) 肿瘤坏死因子(TNF) 3-磷酸甘油醛脱氢酶(GAPDH) 负压伤口疗法(NPWT) 血管内皮生长因子(VEGF) 苏木精-伊红(HE) 动脉血二氧化碳分压(PaCO2) 负压封闭引流(VSD) 所有作者均声明不存在利益冲突广告目次深圳市源兴医药股份有限公司 ………………………………………………………………………………………… 插页3美纳里尼(中国)投资有限公司 ………………………………………………………………………………………… 插页5上海铠唏尔医疗器械贸易有限公司 …………………………………………………………………………………… 插页6南海朗肽制药有限公司 ………………………………………………………………………………………… 对中文目次1江西省科星生物工程有限公司 ………………………………………………………………………………… 对中文目次2上海腾瑞制药股份有限公司 …………………………………………………………………………………… 对英文目次1保赫曼(上海)贸易有限公司 …………………………………………………………………………………… 对英文目次2浙江医学科技开发有限公司 …………………………………………………………………………………………… 插页7苏州汇涵医用科技发展有限公司 ……………………………………………………………………………………… 插页8苏州爱得科技发展股份有限公司 ……………………………………………………………………………………… 对正文珠海亿胜生物制药有限公司 ……………………………………………………………………………………………… 封三武汉维斯第医用科技股份有限公司 ……………………………………………………………………………………… 封底 -
参考文献
(27) [1] BurgessJL,WyantWA,Abdo AbujamraB,et al.Diabetic wound-healing science[J].Medicina (Kaunas),2021,57(10):1072. DOI: 10.3390/medicina57101072. [2] RatajczakJ,WysoczynskiM,HayekF,et al.Membrane-derived microvesicles: important and underappreciated mediators of cell-to-cell communication[J].Leukemia,2006,20(9):1487-1495.DOI: 10.1038/sj.leu.2404296. [3] FerreiraADF, GomesDA. Stem cell extracellular vesicles in skin repair[J].Bioengineering (Basel),2018,6(1):4. DOI: 10.3390/bioengineering6010004. [4] van NielG, D'AngeloG, RaposoG.Shedding light on the cell biology of extracellular vesicles[J].Nat Rev Mol Cell Biol,2018,19(4):213-228. DOI: 10.1038/nrm.2017.125. [5] ZhangW,BaiX,ZhaoB,et al.Cell-free therapy based on adipose tissue stem cell-derived exosomes promotes wound healing via the PI3K/Akt signaling pathway[J].Exp Cell Res,2018,370(2):333-342. DOI: 10.1016/j.yexcr.2018.06.035. [6] ZhangJ,ChenC,HuB,et al.Exosomes derived from human endothelial progenitor cells accelerate cutaneous wound healing by promoting angiogenesis through Erk1/2 signaling[J].Int J Biol Sci,2016,12(12):1472-1487.DOI: 10.7150/ijbs.15514. [7] LiX, LiuL, YangJ, et al. Exosome derived from human umbilical cord mesenchymal stem cell mediates miR-181c attenuating burn-induced excessive inflammation[J]. EBioMedicine, 2016,8:72-82. DOI: 10.1016/j.ebiom.2016.04.030. [8] ZhaoB,LiX,ShiX,et al.Exosomal microRNAs derived from human amniotic epithelial cells accelerate wound healing by promoting the proliferation and migration of fibroblasts[J].Stem Cells Int,2018,2018:5420463.DOI: 10.1155/2018/5420463. [9] ZhengY,ZhengS,FanX,et al.Amniotic epithelial cells accelerate diabetic wound healing by modulating inflammation and promoting neovascularization[J].Stem Cells Int,2018,2018:1082076.DOI: 10.1155/2018/1082076. [10] ZhengY,JiS,WuH,et al.Topical administration of cryopreserved living micronized amnion accelerates wound healing in diabetic mice by modulating local microenvironment[J].Biomaterials,2017,113:56-67.DOI: 10.1016/j.biomaterials.2016.10.031. [11] WeiP,ZhongC,YangX,et al.Exosomes derived from human amniotic epithelial cells accelerate diabetic wound healing via PI3K-AKT-mTOR-mediated promotion in angiogenesis and fibroblast function[J/OL].Burns Trauma,2020,8:tkaa020[2021-04-24].https://pubmed.ncbi.nlm.nih.gov/32923490/.DOI: 10.1093/burnst/tkaa020. [12] XiaoGY,ChengCC,ChiangYS,et al.Exosomal miR-10a derived from amniotic fluid stem cells preserves ovarian follicles after chemotherapy[J].Sci Rep,2016,6:23120.DOI: 10.1038/srep23120. [13] FortunatoO,GaspariniP,BoeriM,et al.Exo-miRNAs as a new tool for liquid biopsy in lung cancer[J].Cancers (Basel),2019,11(6):888. DOI: 10.3390/cancers11060888. [14] HuY,RaoSS,WangZX,et al.Exosomes from human umbilical cord blood accelerate cutaneous wound healing through miR-21-3p-mediated promotion of angiogenesis and fibroblast function[J].Theranostics,2018,8(1):169-184.DOI: 10.7150/thno.21234. [15] HuangH,CuiW,QiuW,et al.Impaired wound healing results from the dysfunction of the Akt/mTOR pathway in diabetic rats[J].J Dermatol Sci,2015,79(3):241-251.DOI: 10.1016/j.jdermsci.2015.06.002. [16] CastilhoRM,SquarizeCH,GutkindJS.Exploiting PI3K/mTOR signaling to accelerate epithelial wound healing[J].Oral Dis,2013,19(6):551-558.DOI: 10.1111/odi.12070. [17] ShawTJ,MartinP.Wound repair: a showcase for cell plasticity and migration[J].Curr Opin Cell Biol,2016,42:29-37.DOI: 10.1016/j.ceb.2016.04.001. [18] HuSC,LanCE.High-glucose environment disturbs the physiologic functions of keratinocytes: focusing on diabetic wound healing[J].J Dermatol Sci,2016,84(2):121-127.DOI: 10.1016/j.jdermsci.2016.07.008. [19] ZhaoB,LiuJQ,YangC,et al.Human amniotic epithelial cells attenuate TGF-β1-induced human dermal fibroblast transformation to myofibroblasts via TGF-β1/Smad3 pathway[J].Cytotherapy,2016,18(8):1012-1024.DOI: 10.1016/j.jcyt.2016.04.009. [20] ZhaoB,LiuJQ,ZhengZ,et al.Human amniotic epithelial stem cells promote wound healing by facilitating migration and proliferation of keratinocytes via ERK, JNK and AKT signaling pathways[J].Cell Tissue Res,2016,365(1):85-99.DOI: 10.1007/s00441-016-2366-1. [21] CamussiG,DeregibusMC,BrunoS,et al.Exosomes/microvesicles as a mechanism of cell-to-cell communication[J].Kidney Int,2010,78(9):838-848.DOI: 10.1038/ki.2010.278. [22] FriedmanRC,FarhKK,BurgeCB,et al.Most mammalian mRNAs are conserved targets of microRNAs[J].Genome Res,2009,19(1):92-105.DOI: 10.1101/gr.082701.108. [23] MengZ,ZhouD,GaoY,et al.miRNA delivery for skin wound healing[J].Adv Drug Deliv Rev,2018,129:308-318.DOI: 10.1016/j.addr.2017.12.011. [24] MulhollandEJ,DunneN,McCarthyHO.MicroRNA as therapeutic targets for chronic wound healing[J].Mol Ther Nucleic Acids,2017,8:46-55.DOI: 10.1016/j.omtn.2017.06.003. [25] FahsF, BiX, YuFS, et al. New insights into microRNAs in skin wound healing[J]. IUBMB Life, 2015,67(12):889-896. DOI: 10.1002/iub.1449. [26] LiD,LiXI,WangA,et al.MicroRNA-31 promotes skin wound healing by enhancing keratinocyte proliferation and migration[J].J Invest Dermatol,2015,135(6):1676-1685.DOI: 10.1038/jid.2015.48. [27] DeppeJ,SteinritzD,SantovitoD,et al.Upregulation of miR-203 and miR-210 affect growth and differentiation of keratinocytes after exposure to sulfur mustard in normoxia and hypoxia[J].Toxicol Lett,2016,244:81-87.DOI: 10.1016/j.toxlet.2015.09.012. -
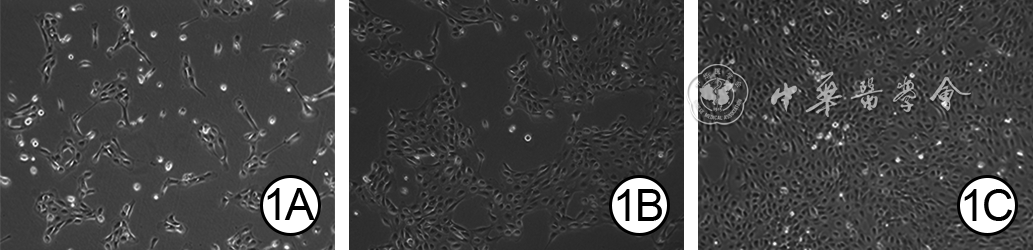
1 原代人羊膜上皮细胞(hAEC)培养第2、4、7天形态观察 倒置荧光显微镜×100。1A.培养第2天,hAEC贴壁生长,形态、大小均一;1B、1C.分别为培养第4、7天,hAEC增殖活跃,细胞密度增加,呈典型的鹅卵石样单层排列
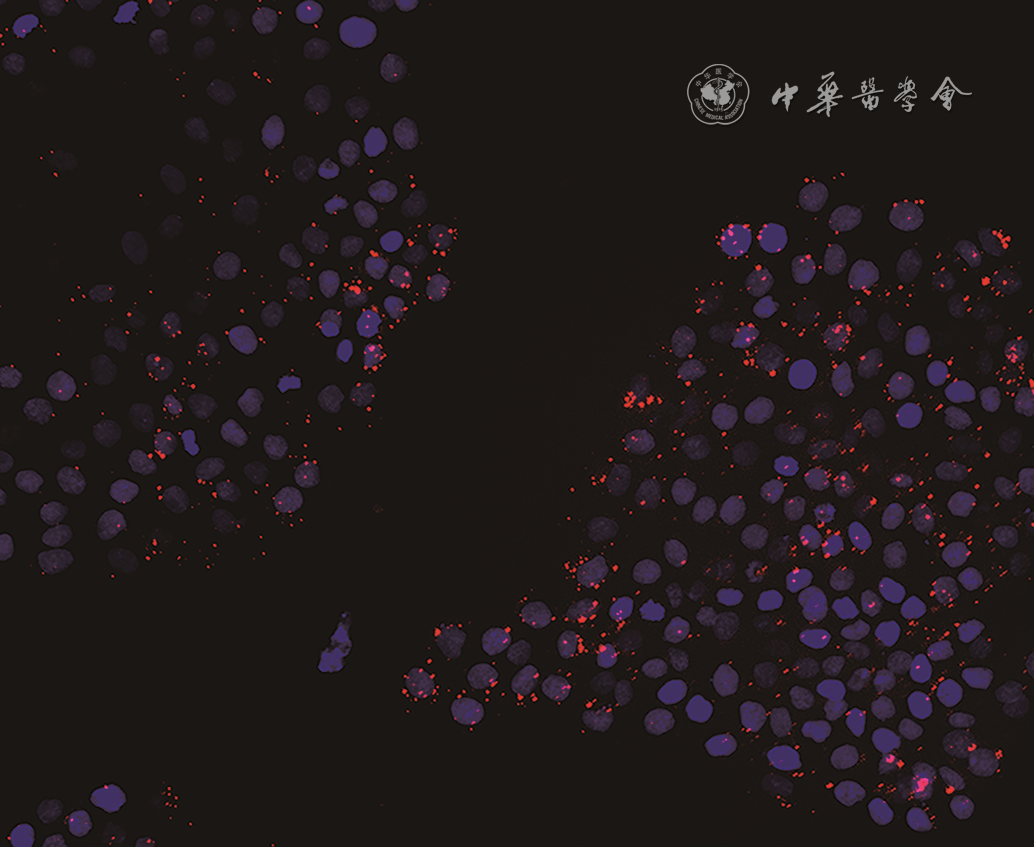
2 人羊膜上皮细胞外泌体(hAEC-Exo)与HaCaT共培养3 h,hAEC-Exo被HaCaT内吞入细胞质,并聚集于细胞核周围 PKH26-4',6-二脒基-2-苯基吲哚×200
注:红色指示外泌体,蓝色指示细胞核
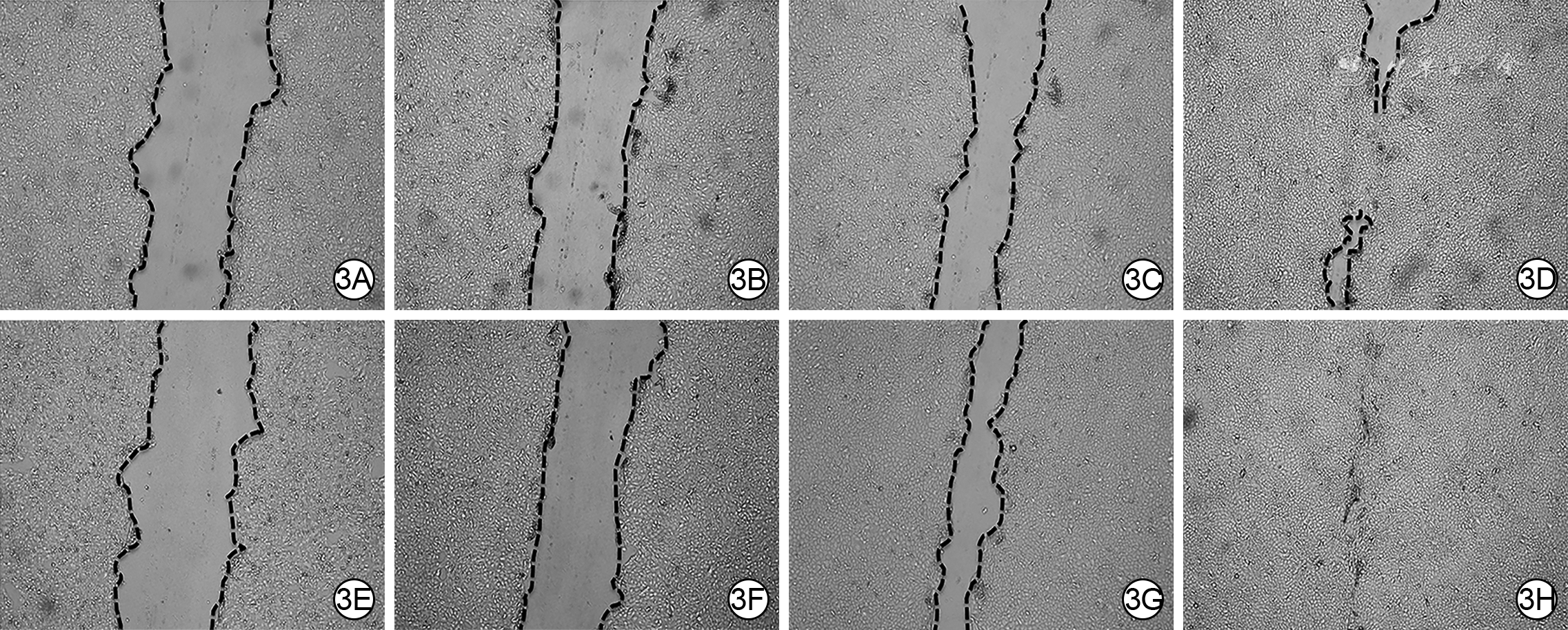
3 2组HaCaT划痕后各时间点划痕面积 倒置荧光显微镜×100。3A、3B、3C、3D.分别为PBS组划痕后0(即刻)、24、48、72 h的划痕面积;3E、3F、3G、3H.分别为hAEC-Exo组划痕后0、24、48、72 h的划痕面积,图3F、3G、3H划痕面积分别明显小于图3B、3C、3D
注:PBS为磷酸盐缓冲液,hAEC-Exo为人羊膜上皮细胞外泌体
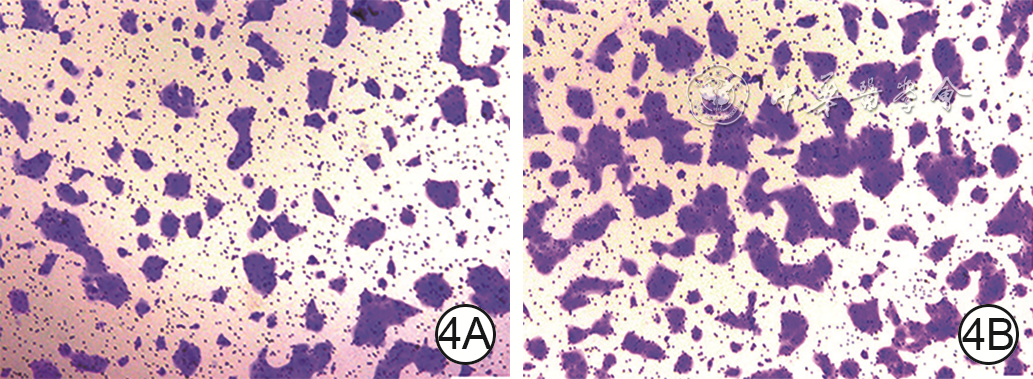
4 Transwell实验观察2组HaCaT培养48 h穿膜迁移情况 苏木精×50。4A、4B.分别为PBS组和hAEC-Exo组,图4B穿膜细胞数明显多于图4A
注:蓝色指示穿膜细胞;PBS为磷酸盐缓冲液,hAEC-Exo为人羊膜上皮细胞外泌体
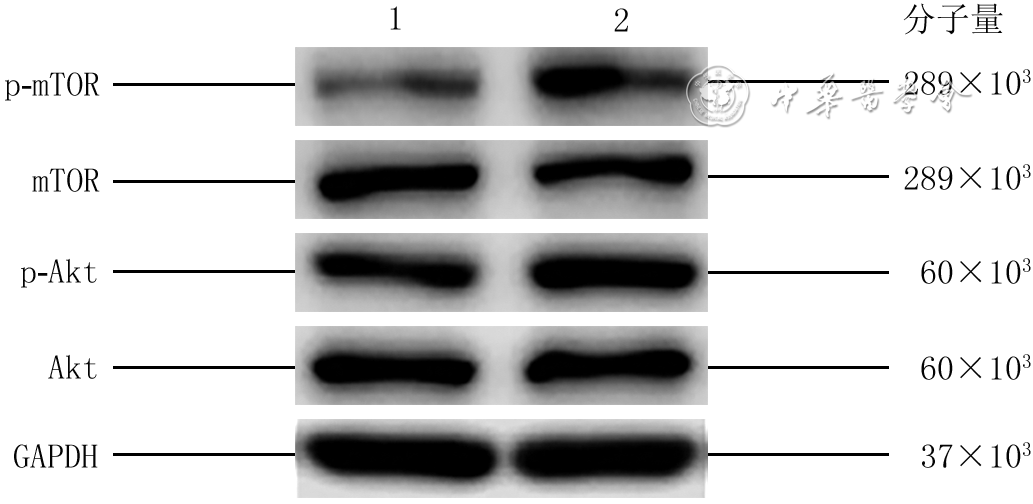
5 蛋白质印迹法检测2组HaCaT培养24 h PI3K-Akt-mTOR通路相关蛋白的表达水平
注:PI3K为磷脂酰肌醇3-激酶,mTOR为哺乳动物雷帕霉素靶蛋白,p-mTOR为磷酸化mTOR,Akt为蛋白激酶B,p-Akt为磷酸化Akt,GAPDH为3-磷酸甘油醛脱氢酶;1、2分别为磷酸盐缓冲液组和人羊膜上皮细胞外泌体组

6 蛋白质印迹法检测3组HaCaT培养24 h PI3K-Akt-mTOR通路相关蛋白的表达量
注:PI3K为磷脂酰肌醇3-激酶,mTOR为哺乳动物雷帕霉素靶蛋白,p-mTOR为磷酸化mTOR,Akt为蛋白激酶B,p-Akt为磷酸化Akt,GAPDH为3-磷酸甘油醛脱氢酶;1、2、3分别为二甲基亚砜(DMSO)+磷酸盐缓冲液组,DMSO+人羊膜上皮细胞外泌体(hAEC-Exo)组,LY294002+hAEC-Exo组

7 3组HaCaT划痕后各时间点划痕面积。7A、7B、7C、7D.分别为DMSO+PBS组划痕后0(即刻)、24、48、72 h的划痕面积;7E、7F、7G、7H.分别为DMSO+hAEC-Exo组划痕后0、24、48、72 h的划痕面积,图7F、7G、7H划痕面积明显小于图7B、7C、7D;7I、7J、7K、7L.分别为LY294002+hAEC-Exo组划痕后0、24、48、72 h的划痕面积,图7J、7K、7L划痕面积明显大于图7F、7G、7H
注:DMSO为二甲基亚砜,PBS为磷酸盐缓冲液,hAEC-Exo为人羊膜上皮细胞外泌体
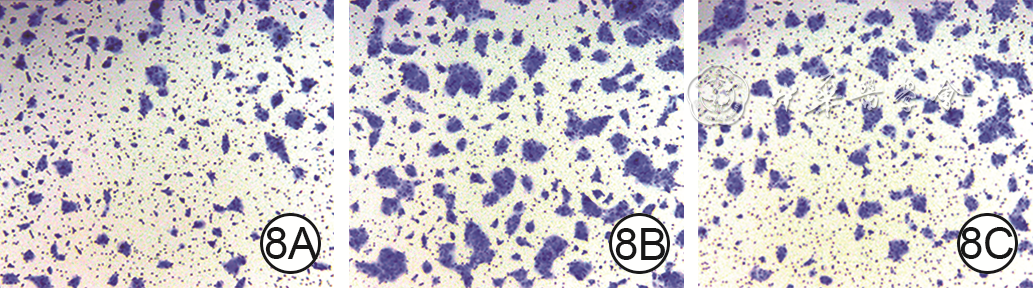
8 Transwell实验观察3组HaCaT培养48 h穿膜迁移情况 苏木精×50。8A、8B、8C.分别为DMSO+PBS组、DMSO+hAEC-Exo组和LY294002+hAEC-Exo组,图8B穿膜迁移细胞数明显多于图8A、8C
注:图中蓝色指示穿膜细胞;DMSO为二甲基亚砜, PBS为磷酸盐缓冲液,hAEC-Exo为人羊膜上皮细胞外泌体
表 1 2组HaCaT培养各时间点增殖活力比较(
组别 样本数 12 h 24 h 36 h 48 h 60 h PBS组 3 0.273±0.025 0.386±0.028 0.494±0.030 0.789±0.060 0.943±0.040 hAEC-Exo组 3 0.404±0.030 0.770±0.041 0.923±0.026 1.161±0.042 1.471±0.038 t值 3.691 10.861 12.121 10.531 14.931 P值 <0.01 <0.01 <0.01 <0.01 <0.01 注:PBS为磷酸盐缓冲液,hAEC-Exo为人羊膜上皮细胞外泌体;处理因素主效应,F=783.2,P<0.01;时间因素主效应,F=494.7,P<0.01;两者交互作用,F=27.0,P<0.01  下载: 导出CSV
下载: 导出CSV
表2 2组HaCaT划痕后各时间点划痕愈合率比较
表2. (%,x¯±s)
组别 样本数 24 h 48 h 72 h PBS组 3 22.6±2.4 42.8±4.0 74.5±4.5 hAEC-Exo组 3 34.7±4.9 66.1±4.8 94.4±4.1 t值 3.342 6.427 5.485 P值 <0.05 <0.01 <0.01 注:PBS为磷酸盐缓冲液,hAEC-Exo为人羊膜上皮细胞外泌体;处理因素主效应,F=692.7,P<0.01;时间因素主效应,F=25.4,P<0.01;两者交互作用,F=14.4,P<0.01
下载: 导出CSV
表3 2组HaCaT培养24 h PI3K-Akt-mTOR通路相关蛋白的表达量比较(
组别 样本数 mTOR p-mTOR Akt p-Akt PBS组 3 0.517±0.045 0.267±0.066 0.823±0.068 0.333±0.050 hAEC-Exo组 3 0.537±0.049 0.437±0.046 0.787±0.025 0.610±0.029 t值 0.404 4.240 0.740 5.586 P值 >0.05 <0.01 >0.05 <0.01 注:PBS为磷酸盐缓冲液,hAEC-Exo为人羊膜上皮细胞外泌体;PI3K为磷脂酰肌醇3-激酶,Akt为蛋白激酶B,p-Akt为磷酸化Akt,mTOR为哺乳动物雷帕霉素靶蛋白,p-mTOR为磷酸化mTOR
下载: 导出CSV
表4 3组HaCaT中培养24 h PI3K-Akt-mTOR通路相关蛋白的表达量比较(
组别 样本数 mTOR p-mTOR Akt p-Akt DMSO+PBS组 3 0.716±0.017 0.395±0.038 0.859±0.033 0.228±0.028 DMSO+hAEC-Exo组 3 0.716±0.026 0.600±0.054 0.847±0.029 0.505±0.034 LY294002+hAEC-Exo组 3 0.699±0.027 0.433±0.029 0.865±0.039 0.374±0.033 F值 0.4 13.8 0.1 38.3 P值 >0.05 <0.01 >0.05 <0.01 t1值 0.024 6.155 0.353 8.338 P1值 >0.05 <0.01 >0.05 <0.01 t2值 0.501 5.030 0.517 3.960 P2值 >0.05 <0.01 >0.05 <0.01 注:DMSO为二甲基亚砜,PBS为磷酸盐缓冲液,hAEC-Exo为人羊膜上皮细胞外泌体,PI3K为磷脂酰肌醇3-激酶,Akt为蛋白激酶B,p-Akt为磷酸化Akt,mTOR为哺乳动物雷帕霉素靶蛋白,p-mTOR为磷酸化mTOR;t1值、P1值,t2值、P2值分别DMSO+hAEC-Exo组与DMSO+PBS组、LY294002+hAEC-Exo组比较所得
下载: 导出CSV
表5 3组HaCaT培养各时间点增殖活力比较(
组别 样本数 12 h 24 h 36 h 48 h 60 h DMSO+PBS组 3 0.46±0.04 0.69±0.07 0.98±0.08 1.16±0.08 1.26±0.11 DMSO+hAEC-Exo组 3 0.78±0.05 1.23±0.07 1.60±0.09 1.86±0.09 2.03±0.08 LY294002+hAEC-Exo组 3 0.59±0.03 0.96±0.09 1.20±0.08 1.39±0.08 1.55±0.10 t1值 4.376 7.398 8.488 9.766 10.730 P1值 <0.01 <0.01 <0.01 <0.01 <0.01 t2值 2.691 3.639 5.447 6.605 6.693 P2值 >0.05 <0.05 <0.01 <0.01 <0.01 注:DMSO为二甲基亚砜,PBS为磷酸盐缓冲液,hAEC-Exo为人羊膜上皮细胞外泌体;处理因素主效应,F=141.3,P<0.01;时间因素主效应,F=279.1,P<0.01;两者交互作用,F=8.2,P<0.01;t1值、P1值,t2值、P2值分别DMSO+hAEC-Exo组与DMSO+PBS组、LY294002+hAEC-Exo组比较所得
下载: 导出CSV
表6 3组HaCaT培养各时间点划痕愈合率比较(%,
组别 样本数 24 h 48 h 72 h DMSO+PBS组 3 11.2±1.5 35.2±4.4 71.2±3.4 DMSO+hAEC-Exo组 3 23.0±2.0 54.0±3.9 94.0±2.9 LY294002+hAEC-Exo组 3 11.8±1.8 40.1±2.5 82.0±2.9 t1值 4.003 6.349 7.714 P1值 <0.01 <0.01 <0.01 t2值 3.805 4.676 4.067 P2值 <0.05 <0.01 <0.01 注:DMSO为二甲基亚砜,PBS为磷酸盐缓冲液,hAEC-Exo为人羊膜上皮细胞外泌体;处理因素主效应,F=779.4,P<0.01;时间因素主效应,F=57.1,P<0.01;两者交互作用,F=2.2,P=0.11;t1值、P1值,t2值、P2值分别DMSO+hAEC-Exo组与DMSO+PBS组、LY294002+hAEC-Exo组比较所得
下载: 导出CSV
-

 下载:
下载:
计量
- 文章访问数: 477
- HTML全文浏览量: 115
- PDF下载量: 48
- 被引次数: 0
