



Effects and cell signaling mechanism of glutamine on rat cardiomyocytes intervened with serum from burned rat
-
摘要:
目的 探讨谷氨酰胺对经烧伤大鼠血清(以下简称烧伤血清)干预的大鼠心肌细胞的作用及其细胞信号机制。 方法 采用实验研究方法。取10只7~8个月龄雌雄各半Wistar大鼠制备正常大鼠血清(以下简称正常血清),另取20只7~8个月龄雌雄各半Wistar大鼠造成30%体表总面积Ⅲ度烧伤后制备烧伤血清,从180只1~3 d龄雌雄不拘Wistar大鼠心尖组织中分离培养原代心肌细胞用于后续实验。按随机数字表法(分组方法下同)将细胞分为正常血清组、烧伤血清组,加入相应血清培养。分别于培养1、3、6、9、12 h,采用锥虫蓝实验检测细胞存活率。将细胞分为单纯烧伤血清组、烧伤血清+4 mmol/L谷氨酰胺组、烧伤血清+8 mmol/L谷氨酰胺组、烧伤血清+12 mmol/L谷氨酰胺组、烧伤血清+16 mmol/L谷氨酰胺组、烧伤血清+20 mmol/L谷氨酰胺组,采用单纯烧伤血清或烧伤血清+相应终物质的量浓度谷氨酰胺处理,培养前实验筛选出的干预时间,同前检测细胞存活率。将细胞分为正常血清组、单纯烧伤血清组、烧伤血清+12 mmol/L谷氨酰胺组、烧伤血清+16 mmol/L谷氨酰胺组、烧伤血清+20 mmol/L谷氨酰胺组,同前处理后采用蛋白质印迹法检测培养30 min哺乳动物雷帕霉素靶蛋白复合物1(mTORC1)、p70核糖体蛋白S6激酶(p70 S6K)和真核翻译起始因子4E结合蛋白1 (4E-BP1)磷酸化水平。将细胞分为正常血清组、单纯烧伤血清组、烧伤血清+12 mmol/L谷氨酰胺组、烧伤血清+12 mmol/L谷氨酰胺+25 ng/mL雷帕霉素组,行相应处理后,分别于培养1、3、6 h,采用免疫荧光法检测热休克蛋白70(HSP70)和金属硫蛋白(MT)表达并观测微管形态。各组各时间点各指标样本数均为10。对数据行析因设计方差分析、单因素方差分析、LSD-t检验、LSD检验及Bonferroni校正。 结果 培养1、3、6、9、12 h,烧伤血清组细胞存活率均明显低于正常血清组(t=4.950、16.752、35.484、34.428、27.781,P<0.01)。与组内培养1 h比,烧伤血清组培养3、6、9、12 h细胞存活率明显降低(P<0.05);与组内培养3 h比较,烧伤血清组培养6、9、12 h细胞存活率明显降低(P<0.05);与组内培养6、9 h比较,烧伤血清组培养12 h细胞存活率明显降低(P<0.05);烧伤血清组培养6、9 h细胞存活率比较,差异无统计学意义(P>0.05)。因此选择培养6 h作为后续烧伤血清干预时间。培养6 h,与单纯烧伤血清组比较,烧伤血清+4 mmol/L谷氨酰胺组、烧伤血清+8 mmol/L谷氨酰胺组、烧伤血清+12 mmol/L谷氨酰胺组、烧伤血清+16 mmol/L谷氨酰胺组、烧伤血清+20 mmol/L谷氨酰胺组细胞存活率均明显升高(P<0.01)。烧伤血清+12 mmol/L谷氨酰胺组与烧伤血清+16 mmol/L谷氨酰胺组细胞存活率相近(P>0.05),烧伤血清+16 mmol/L谷氨酰胺组与烧伤血清+20 mmol/L谷氨酰胺组细胞存活率相近(P>0.05)。因此选择12、16、20 mmol/L作为后续谷氨酰胺的干预浓度。培养30 min,正常血清组、单纯烧伤血清组、烧伤血清+12 mmol/L谷氨酰胺组、烧伤血清+16 mmol/L谷氨酰胺组、烧伤血清+20 mmol/L谷氨酰胺组细胞mTORC1、p70 S6K、4E-BP1磷酸化水平分别为1.001±0.042、0.510±0.024、0.876±0.022、0.836±0.074、0.856±0.041,1.00±0.11、0.38±0.09、0.95±0.13、0.96±0.13、0.89±0.24,1.00±0.07、0.29±0.08、0.87±0.27、0.68±0.08、0.60±0.21。与正常血清组比较,其余4个烧伤血清组细胞mTORC1、p70 S6K、4E-BP1磷酸化水平均明显降低(P<0.01) 。 与单纯烧伤血清组比较,其余3个烧伤血清组细胞mTORC1、p70 S6K、4E-BP1磷酸化水平均明显升高(P<0.01)。烧伤血清+12 mmol/L谷氨酰胺组细胞4E-BP1磷酸化水平明显高于烧伤血清+16 mmol/L谷氨酰胺组和烧伤血清+20 mmol/L谷氨酰胺组(P<0.05)。单纯烧伤血清组细胞培养1 h MT表达明显低于正常血清组(P<0.05),其余时间点MT表达明显高于正常血清组(P<0.01);培养1、3、6 h,单纯烧伤血清组细胞HSP70表达明显高于正常血清组(P<0.05),烧伤血清+12 mmol/L谷氨酰胺组细胞HSP70和MT表达均明显高于单纯烧伤血清组(P<0.05),烧伤血清+12 mmol/L谷氨酰胺+25 ng/mL雷帕霉素组细胞HSP70和MT表达均明显低于烧伤血清+12 mmol/L谷氨酰胺组(P<0.01)。正常血清组细胞培养1、3、6 h微管结构完整,呈网格排列,染色均匀。单纯烧伤血清组细胞培养1 h部分微管出现断裂,部分网格排列不齐;培养3 h靠近细胞核微管结构清晰,细胞核远端微管模糊不清;培养6 h微管结构模糊不清。烧伤血清+12 mmol/L谷氨酰胺组细胞培养各时间点微管破坏程度较单纯烧伤血清组轻。烧伤血清+12 mmol/L谷氨酰胺+25 ng/mL雷帕霉素组细胞培养各时间点微管形态与单纯烧伤血清组相近。 结论 烧伤血清可导致大鼠心肌细胞受损,细胞存活率明显下降。谷氨酰胺通过调控mTOR/p70 S6K/4E-BP1信号通路,促进心肌细胞HSP70和MT表达,稳定微管结构,从而发挥其细胞保护作用。 -
关键词:
- 烧伤 /
- 谷氨酰胺 /
- 肌细胞,心脏 /
- 哺乳动物雷帕霉素靶蛋白信号通路
Abstract:Objective To investigate the effects and cell signaling mechanism of glutamine on rat cardiomyocytes intervened with serum from burned rat (hereinafter referred to as burn serum). Methods The experimental research method was applied. Ten gender equally distributed Wistar rats aged 7-8 months were taken to prepare normal rat serum (hereinafter referred to as normal serum), another twenty gender equally distributed Wistar rats aged 7-8 months were taken to prepare burn serum after full- thickness burn injury of 30% total body surface area, and primary cardiomyocytes were isolated and cultured from 180 Wistar rats aged 1-3 days by either gender and used in the following experiments. The cells were divided into normal serum group and burn serum group according to the random number table (the same grouping method below) and cultured with the corresponding serum. At post culture hour (PCH) 1, 3, 6, 9, and 12, trypanosoma blue test was used to detect the cell survival rate. The cells were divided into burn serum alone group, burn serum+4 mmol/L glutamine group, burn serum+8 mmol/L glutamine group, burn serum+12 mmol/L glutamine group, burn serum+16 mmol/L glutamine group, and burn serum+20 mmol/L glutamine group, which were treated with burn serum alone or burn serum added with the corresponding final molarity of glutamine and cultured for the time screened in the experiment before, and then the cell survival rate was detected as before. The cells were divided into normal serum group, burn serum alone group, burn serum+12 mmol/L glutamine group, burn serum+16 mmol/L glutamine group, and burn serum+20 mmol/L glutamine group and treated the same as before. After 30 min of culture, phosphorylation levels of mammalian target of rapamycin complex 1 (mTORC1), p70 ribosomal protein S6 kinase (p70 S6K), and eukaryotic translation initiation factor 4E-binding protein 1 (4E-BP1) were detected by Western blotting. Cells were divided into normal serum group, burn serum alone group, burn serum+12 mmol/L glutamine group, burn serum+12 mmol/L glutamine+25 ng/mL rapamycin group, and treated correspondingly. At PCH 1, 3, and 6, the expressions of heat shock protein 70 (HSP70) and metallothionein (MT), and the morphology of microtubule were determined with immunofluorescence method. The sample numbers in each index at each time point in each group were all 10. Data were statistically analyzed with analysis of variance for factorial design, one-way analysis of variance, least significant difference t test, least significant difference test, and Bonferroni correction. Results At PCH 1, 3, 6, 9, and 12, the cell survival rates in burn serum group were significantly lower than those in normal serum group (t=4.950, 16.752, 35.484, 34.428, 27.781, P<0.01). Compared within the group at PCH 1, the cell survival rate was significantly decreased in burn serum group at PCH 3, 6, 9, and 12 (P<0.05). Compared within the group at PCH 3, the cell survival rate was significantly decreased in burn serum group at PCH 6, 9, and 12 (P<0.05). Compared within the group at PCH 6 and 9, the cell survival rate was significantly decreased in burn serum group at PCH 12 (P<0.05). There were no statistically significant differences in the cell survival rates in burn serum group between PCH 6 and 9 (P>0.05). Thus PCH 6 was selected as the subsequent intervention time of burn serum. At PCH 6, compared with burn serum alone group, the cell survival rates in burn serum+4 mmol/L glutamine group, burn serum+8 mmol/L glutamine group, burn serum+12 mmol/L glutamine group, burn serum+16 mmol/L glutamine group, and burn serum+20 mmol/L glutamine group were significantly increased (P<0.01). There were no statistically significant differences in cell survival rates between burn serum+12 mmol/L glutamine group and burn serum+16 mmol/L glutamine group (P>0.05). There were no statistically significant differences in cell survival rates in burn serum+16 mmol/L glutamine group and burn serum+20 mmol/L glutamine group (P>0.05). Thus 12, 16, and 20 mmol/L were selected as the subsequent intervention concentrations of glutamine. After 30 min of culture, the phosphorylation levels of mTORC1, p70 S6K, and 4E-BP1 of cells were respectively 1.001±0.042, 0.510±0.024, 0.876±0.022, 0.836±0.074, 0.856±0.041, 1.00±0.11, 0.38±0.09, 0.95±0.13, 0.96±0.13, 0.89±0.24, 1.00±0.07, 0.29±0.08, 0.87±0.27, 0.68±0.08, 0.60±0.21 in normal serum group, burn serum alone group, burn serum+12 mmol/L glutamine group, burn serum+16 mmol/L glutamine group, and burn serum+20 mmol/L glutamine group. Compared with normal serum group, the phosphorylation levels of mTORC1, p70 S6K, and 4E-BP1 of cells were significantly decreased in the other 4 burn serum groups (P<0.01). Compared with those of burn serum alone group, the phosphorylation levels of mTORC1, p70 S6K, and 4E-BP1 of cells were significantly increased in the other 3 burn serum groups (P<0.01). The phosphorylation level of 4E-BP1 of cells in burn serum+12 mmol/L glutamine group was significantly higher than the levels in burn serum+16 mmol/L glutamine group and burn serum+20 mmol/L glutamine group (P<0.05). The expression of MT of cells in burn serum alone group was significantly lower than that in normal serum group at PCH 1 (P<0.05), while the expressions of MT of cells in burn serum alone group were significantly higher than those in normal serum group at the other time points (P<0.05). At PCH 1, 3, and 6, the expressions of HSP70 of cells in burn serum alone group were significantly higher than those in normal serum group (P<0.05), the expressions of HSP70 and MT of cells in burn serum+12 mmol/L glutamine group were significantly higher than those in burn serum alone group (P<0.05), and the expressions of HSP70 and MT of cells in burn serum+12 mmol/L glutamine+25 ng/mL rapamycin group were significantly lower than those in burn serum+12 mmol/L glutamine group (P<0.01). The microtubular structures were intact, displaying grid alinement and uniform staining in cells of normal serum group at PCH 1, 3, and 6. In burn serum alone group, some microtubules showed fracture and irregular grid arrangement at PCH 1; the microtubular structures near the nucleus were clear, while the microtubules at the distal end of the nucleus were blurry at PCH 3; the microtubular structures were blurry at PCH 6. The microtubular damage of cells was alleviated in burn serum+12 mmol/L glutamine group as compared with that in burn serum alone group at each time point of culture. The morphology of microtubules of cells in burn serum+12 mmol/L glutamine+25 ng/mL rapamycin group at each time point of culture was similar to that of burn serum alone group. Conclusions The burn serum can lead to damages to cardiomyocytes and significant decrease of cell survival rate in rats. Glutamine can exert cell protective function through the regulation of mTOR/p70 S6K/4E-BP1 signaling pathway, thus promoting the expressions of HSP70 and MT and stabilizing the microtubule structures. -
Key words:
- Burns /
- Glutamine /
- Myocytes, cardiac /
- Mammalian target of rapamycin signaling pathway
-
所有作者均声明不存在利益冲突
-
参考文献
(33) [1] 彭曦. 烧伤高代谢机制的再认识及调控策略[J].中华烧伤杂志,2013,29(2):139-143. DOI: 10.3760/cma.j.issn.1009-2587.2013.02.012. [2] 黄跃生. 自噬与严重烧伤后心肌缺血缺氧损害[J].中华烧伤杂志,2018,34(1):3-7. DOI: 10.3760/cma.j.issn.1009-2587.2018.01.002. [3] VermaA, SumiS, SeerviM. Heat shock proteins-driven stress granule dynamics: yet another avenue for cell survival[J]. Apoptosis, 2021,26(7/8):371-384. DOI: 10.1007/s10495-021-01678-w. [4] KurashovaNA, MadaevaIM, KolesnikovaLI. Expression of heat shock proteins HSP70 under oxidative stress[J]. Adv Gerontol, 2019,32(4):502-508. [5] 吕尚军, 张勇, 孙勇, 等. 甘氨酰谷氨酰胺二肽对烧伤大鼠心功能的保护作用[J].中华烧伤杂志,2007,23(4):244-248. DOI: 10.3760/cma.j.issn.1009-2587.2007.04.002. [6] 彭曦. 重视谷氨酰胺在烧伤临床的规范应用[J].肠外与肠内营养,2021,28(1):1-4. DOI: 10.16151/j.1007-810x.2021.01.001. [7] KimuraN, TokunagaC, DalalS, et al. A possible linkage between AMP-activated protein kinase (AMPK) and mammalian target of rapamycin (mTOR) signalling pathway[J]. Genes Cells, 2003,8(1):65-79. DOI: 10.1046/j.1365-2443.2003.00615.x. [8] SaraviaJ, RaynorJL, ChapmanNM, et al. Signaling networks in immunometabolism[J]. Cell Res, 2020,30(4):328-342. DOI: 10.1038/s41422-020-0301-1. [9] RindomE, KristensenAM, OvergaardK, et al. Estimation of p70S6K Thr389 and 4E-BP1 Thr37/46 phosphorylation support dependency of tension per se in a dose-response relationship for downstream mTORC1 signalling[J]. Acta Physiol (Oxf), 2020,229(1):e13426. DOI: 10.1111/apha.13426. [10] HuangHL, LongLY, ZhouPP, et al. mTOR signaling at the crossroads of environmental signals and T-cell fate decisions[J]. Immunol Rev, 2020,295(1):15-38. DOI: 10.1111/imr.12845. [11] GonzálezA, HallMN, LinSC, et al. AMPK and TOR: the Yin and Yang of cellular nutrient sensing and growth control[J]. Cell Metab, 2020,31(3):472-492. DOI: 10.1016/j.cmet.2020.01.015. [12] MacFieJ, McNaughtC. Glutamine and gut barrier function[J]. Nutrition, 2002,18(5):433-434. DOI: 10.1016/s0899-9007(02)00766-9. [13] YangYJ, LiuMM, ZhangY, et al. Effectiveness and mechanism study of glutamine on alleviating hypermetabolism in burned rats[J]. Nutrition, 2020,79/80:110934. DOI: 10.1016/j.nut.2020.110934. [14] YuHD, ZhangYM, ZhangZY, et al. Towards identification of molecular mechanism in which the overexpression of wheat cytosolic and plastid glutamine synthetases in tobacco enhanced drought tolerance[J]. Plant Physiol Biochem, 2020,151:608-620. DOI: 10.1016/j.plaphy.2020.04.013. [15] XiaY, WenHY, YoungME, et al. Mammalian target of rapamycin and protein kinase A signaling mediate the cardiac transcriptional response to glutamine[J]. J Biol Chem, 2003,278(15):13143-13150. DOI: 10.1074/jbc.M208500200. [16] ShawRJ. mTOR signaling: RAG GTPases transmit the amino acid signal[J]. Trends Biochem Sci, 2008,33(12):565-568. DOI: 10.1016/j.tibs.2008.09.005. [17] KameiY, HatazawaY, UchitomiR, et al. Regulation of skeletal muscle function by amino acids[J]. Nutrients, 2020,12(1):261.DOI: 10.3390/nu12010261. [18] LaplanteM, SabatiniDM. mTOR signaling in growth control and disease[J]. Cell, 2012,149(2):274-293. DOI: 10.1016/j.cell.2012.03.017. [19] TafurL, KefauverJ, LoewithR. Structural insights into TOR signaling[J]. Genes (Basel), 2020,11(8):885.DOI: 10.3390/genes11080885. [20] SzwedA, KimE, JacintoE. Regulation and metabolic functions of mTORC1 and mTORC2[J]. Physiol Rev, 2021,101(3):1371- 1426. DOI: 10.1152/physrev.00026.2020. [21] SaxtonRA, SabatiniDM. mTOR signaling in growth, metabolism, and disease[J]. Cell, 2017,168(6):960-976. DOI: 10.1016/j.cell.2017.02.004. [22] ParkJH, LeeG, BlenisJ. Structural insights into the activation of mTORC1 on the lysosomal surface[J]. Trends Biochem Sci, 2020,45(5):367-369. DOI: 10.1016/j.tibs.2020.02.004. [23] Francois-VaughanH, AdebayoAO, BrilliantKE, et al. Persistent effect of mTOR inhibition on preneoplastic foci progression and gene expression in a rat model of hepatocellular carcinoma[J]. Carcinogenesis, 2016,37(4):408-419. DOI: 10.1093/carcin/bgw016. [24] YangHJ, RudgeDG, KoosJD, et al. mTOR kinase structure, mechanism and regulation[J]. Nature, 2013,497(7448):217-223. DOI: 10.1038/nature12122. [25] ZhuJ, WangYF, ChaiXM, et al. Exogenous NADPH ameliorates myocardial ischemia-reperfusion injury in rats through activating AMPK/mTOR pathway[J]. Acta Pharmacol Sin, 2020,41(4):535-545. DOI: 10.1038/s41401-019-0301-1. [26] ItoN, RueggUT, TakedaS. ATP-induced increase in intracellular calcium levels and subsequent activation of mTOR as regulators of skeletal muscle hypertrophy[J]. Int J Mol Sci, 2018,19(9):2804.DOI: 10.3390/ijms19092804. [27] ParrottaL, CrestiM, CaiG. Heat-shock protein 70 binds microtubules and interacts with kinesin in tobacco pollen tubes[J]. Cytoskeleton (Hoboken), 2013,70(9):522-537. DOI: 10.1002/cm.21134. [28] YuanAT, KorkolaNC, WongDL, et al. Metallothionein Cd4S11 cluster formation dominates in the protection of carbonic anhydrase[J]. Metallomics, 2020,12(5):767-783. DOI: 10.1039/d0mt00023j. [29] YangLF, MaJP, TanY, et al. Cardiac-specific overexpression of metallothionein attenuates L-NAME-induced myocardial contractile anomalies and apoptosis[J]. J Cell Mol Med, 2019,23(7):4640-4652. DOI: 10.1111/jcmm.14375. [30] 党永明, 房亚东, 胡炯宇, 等.成体大鼠心肌细胞微管解聚对线粒体分布及能量代谢的影响[J] . 中华烧伤杂志,2010,26(1): 18-22. DOI: 10.3760/cma.j.issn.1009-2587.2010.01.006. [31] XiangF, MaSY, LvYL, et al. Tumor necrosis factor receptor- associated protein 1 regulates hypoxia-induced apoptosis through a mitochondria-dependent pathway mediated by cytochrome c oxidase subunit Ⅱ[J/OL]. Burns Trauma, 2019,7:16[2021-06-01]. https://pubmed.ncbi.nlm.nih.gov/31143823/.DOI: 10.1186/s41038-019- 0154-3. [32] 滕苗, 黄跃生, 郑霁, 等. 微管干预剂对大鼠缺氧心肌细胞能量生成的影响[J].中华烧伤杂志,2007,23(3):164-167. DOI: 10.3760/cma.j.issn.1009-2587.2007.03.002. [33] BuruteM, KapiteinLC. Cellular logistics: unraveling the interplay between microtubule organization and intracellular transport[J]. Annu Rev Cell Dev Biol, 2019,35:29-54. DOI: 10.1146/annurev-cellbio-100818-125149. -
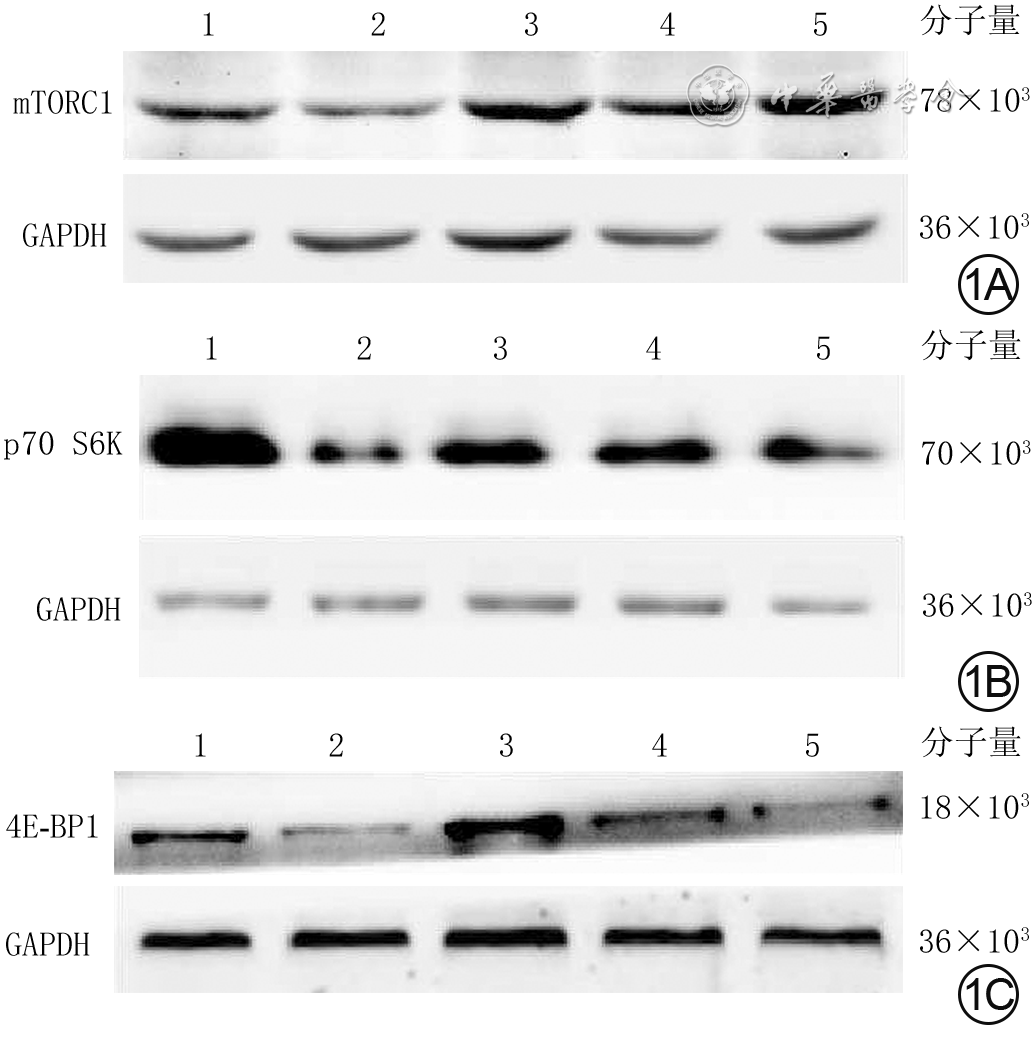
1 蛋白质印迹法检测5组大鼠心肌细胞培养30 min mTORC1、p70 S6K、4E-BP1磷酸化水平。1A.mTORC1磷酸化;1B.p70 S6K磷酸化;1C.4E-BP1磷酸化
注:1、2、3、4、5分别为正常血清组、单纯烧伤血清组、烧伤血清+12 mmol/L谷氨酰胺组、烧伤血清+16 mmol/L谷氨酰胺组、烧伤血清+20 mmol/L谷氨酰胺组;mTORC1为哺乳动物雷帕霉素靶蛋白复合物1,p70 S6K为p70核糖体蛋白S6激酶,4E-BP1为真核翻译起始因子4E结合蛋白1,GAPDH为3-磷酸甘油醛脱氢酶
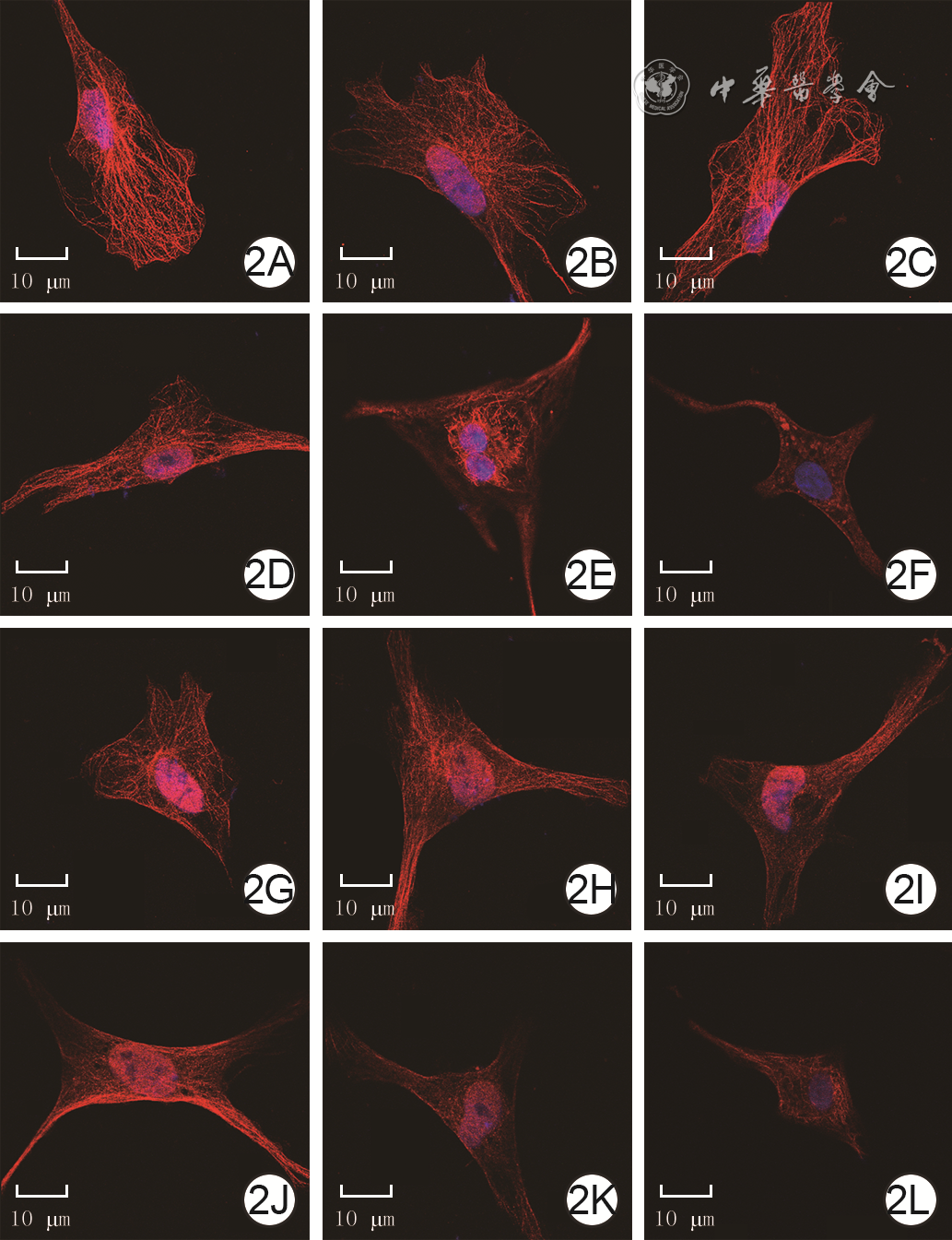
2 免疫荧光法观测4组大鼠心肌细胞培养各时间点微管形态 异硫氰酸荧光素-Hochest 33342×800,图中标尺为10 μm。2A、2B、2C.分别为正常血清组培养1、3、6 h,微管结构完整,呈网格排列,染色均匀,各时间点无明显差异;2D、2E、2F.分别为单纯烧伤血清组培养1、3、6 h,培养1 h部分微管出现断裂且网格排列出现紊乱,培养3 h靠近细胞核微管结构清晰而细胞核远端微管模糊不清,培养6 h微管结构模糊不清且微管染色不均,细胞皱缩;2G、2H、2I.分别为烧伤血清+12 mmol/L谷氨酰胺组培养1、3、6 h,微管破坏程度较图2D、2E、2F有所减轻;2J、2K、2L.分别为烧伤血清+12 mmol/L谷氨酰胺+25 ng/mL雷帕霉素组培养1、3、6 h,微管破坏程度与图2D、2E、2F相近
注:心肌细胞微管阳性染色为红色,细胞核阳性染色为蓝色
表1 2组大鼠心肌细胞培养各时间点存活率比较(%,
组别 样本数 1 h 3 h 6 h 9 h 12 h F值 P值 正常血清组 10 91.4±3.1 88.5±3.5 91.0±3.2 91.4±3.0 89.6±4.0 1.501 >0.05 烧伤血清组 10 83.9±4.2abcd 60.7±3.9bcd 45.3±2.6d 44.9±3.0d 41.8±3.7 240.520 <0.01 t值 4.950 16.752 35.484 34.428 27.781 P值 <0.01 <0.01 <0.01 <0.01 <0.01 注:处理因素主效应,F=1 929.671,P<0.01;时间因素主效应,F=140.609,P<0.01;两者交互作用,F=134.759,P<0.01;F值、P值为组内各时相点总体比较所得;与组内3 h比较,aP<0.05;与组内6 h比较,bP<0.05;与组内9 h比较,cP<0.05;与组内12 h比较,dP<0.05  下载: 导出CSV
下载: 导出CSV
表2 4组大鼠心肌细胞培养各时间点HSP70和MT表达比较(
组别与时间点 样本数 HSP70 MT 正常血清组 30 1 h 3.5±1.2 5.5±1.0 3 h 3.5±0.3 5.5±0.9 6 h 3.5±0.7 5.8±1.1 单纯烧伤血清组 30 1 h 7.5±1.2a 5.2±1.7a 3 h 5.3±1.5a 7.3±1.2a 6 h 4.6±1.6a 8.2±0.9a 烧伤血清+12 mmol/L谷氨酰胺组 30 1 h 8.6±1.6b 10.4±1.0b 3 h 10.2±2.2b 13.0±1.4b 6 h 12.0±1.5b 14.4±1.7b 烧伤血清+12 mmol/L谷氨酰胺+ 25 ng/mL雷帕霉素组 30 1 h 1.5±0.4c 2.4±1.0c 3 h 1.8±0.8c 2.6±0.4c 6 h 2.1±0.9c 2.8±1.1c F1值 79.356 72.462 P1值 <0.01 <0.01 F2值 65.408 181.557 P2值 <0.01 <0.01 F3值 123.931 161.739 P3值 <0.01 <0.01 注:HSP70为热休克蛋白70,MT为金属硫蛋白;各组各时间点样本数均为10;HSP70和MT表达处理因素主效应,F=168.769、335.880,P<0.01;时间因素主效应,F=1.007、26.941,P<0.01;两者交互作用,F=13.433、7.648,P<0.01;F1值、P1值,F2值、P2值,F3值、P3值分别为单纯烧伤血清组、烧伤血清+12 mmol/L谷氨酰胺组、烧伤血清+12 mmol/L谷氨酰胺+25 ng/mL雷帕霉素组培养1、3、6 h各指标总体比较所得;与正常血清组比较,aP<0.05;与单纯烧伤血清组比较,bP<0.05;与烧伤血清+12 mmol/L谷氨酰胺组比较,cP<0.01
下载: 导出CSV
-

计量
- 文章访问数: 972
- HTML全文浏览量: 68
- PDF下载量: 18
- 被引次数: 0
