



Effect of human decidua mesenchymal stem cells-derived exosomes on the function of high glucose-induced senescent human dermal fibroblasts and its possible mechanism
-
摘要:
目的 建立人真皮成纤维细胞(HDF)高糖老化模型,探讨人蜕膜间充质干细胞(dMSC)来源外泌体对高糖老化HDF增殖、迁移、凋亡的影响及其可能机制。 方法 采用实验研究方法。收集2021年1—3月解放军总医院第四医学中心收治的4例男性包茎患者(18~22岁)环切术后废弃包皮组织,分离培养获取原代HDF。取第6代HDF,按照随机数字表法分为低糖组和高糖组,分别采用低糖完全培养基和高糖完全培养基进行每72小时换液、不传代培养,10 d后取细胞,于接种后24 h,采用β-半乳糖苷酶试剂盒检测细胞衰老情况;于接种后48 h,采用蛋白质印迹法检测细胞衰老相关蛋白p16、p53表达情况;于接种后24、48、72 h,采用细胞计数试剂盒8(CCK-8)法检测细胞增殖情况;于接种后48 h,采用脱氧尿嘧啶核苷(EdU)染色法检测细胞增殖情况,采用流式细胞术检测细胞周期及凋亡情况;于接种后24 h,采用Transwell实验测定细胞迁移能力。取人dMSC培养48~72 h,采用差速高速离心法获取其外泌体,采用透射电子显微镜观察dMSC外泌体形态,采用纳米颗粒追踪分析法检测dMSC外泌体的粒径分布,采用蛋白质印迹法检测dMSC外泌体标志蛋白CD9、肿瘤易感基因101(TSG101)的表达。取dMSC外泌体及前述高糖完全培养基诱导老化的HDF共孵育24 h,采用PKH67试剂盒检测细胞摄取外泌体的情况。取前述高糖完全培养基诱导老化的HDF,同前分为单纯高糖组、高糖+低浓度外泌体组、高糖+高浓度外泌体组,分别于高糖完全培养基中加入等体积的磷酸盐缓冲液、终质量浓度为50 μg/mL dMSC外泌体、终质量浓度为100 μg/mL dMSC外泌体进行常规细胞培养。分组后同前于对应时间点采用CCK-8法和EdU染色法、流式细胞术、Transwell实验分别检测细胞增殖、细胞周期和凋亡及细胞迁移情况。根据前述结果,另取经高糖完全培养基诱导老化的HDF,分为单纯高糖组、高糖+高浓度外泌体组并同前处理。分组培养48 h,采用实时荧光定量反转录PCR法检测单纯高糖组和高糖+高浓度外泌体组细胞衰老相关的微小RNA-145-5p(miR-145-5p)、miR-498、miR-503-5p及其靶基因钙/钙调素依赖性蛋白激酶1D(CAMK1D)、人第10号染色体缺失的磷酸酶及张力蛋白同源的基因(PTEN基因)和细胞周期蛋白D1的mRNA表达情况。对数据行析因设计方差分析、单因素方差分析、LSD-t检验和独立样本t检验。 结果 接种后24 h,高糖组HDF β-半乳糖苷酶阳性染色率为(38.4±4.2)%,明显高于低糖组的(16.5±2.2)%(t=4.65,P<0.01)。接种后48 h,高糖组HDF的衰老相关蛋白p16和p53的表达量均明显高于低糖组(t值分别为11.85、3.02,P<0.05或P<0.01)。接种后24、48、72 h,高糖组HDF的增殖活性均明显低于低糖组(t值分别为4.13、9.90、15.12,P<0.01)。接种后48 h,高糖组HDF的EdU阳性染色率明显低于低糖组(t=3.83, P<0.05)。接种后48 h,高糖组HDF周期的G2/M+S亚群在3个亚群(G0/G1、S和G2/M)的占比明显低于低糖组(t=8.74,P<0.01)。接种后24 h,高糖组HDF穿过Transwell滤膜到达下室的细胞数量为(37±6)个,明显少于低糖组的(74±7)个(t=8.42,P<0.01)。接种后48 h,高糖组HDF凋亡率明显高于低糖组(t=8.48,P<0.01)。dMSC外泌体为边缘清晰、大小分布均匀的杯状或者圆形囊泡,粒径基本处于80~200 nm。dMSC外泌体标志性蛋白CD9、TSG101表达均呈阳性。共孵育24 h,外泌体被HDF摄入胞内,主要分布于细胞核周围。分组培养24、48、72 h,高糖+低浓度外泌体组和高糖+高浓度外泌体组HDF增殖活性均明显高于单纯高糖组(t值分别为6.36、6.10、7.76,8.92、12.17、10.74,P<0.01),高糖+高浓度外泌体组HDF增殖活性均明显高于高糖+低浓度外泌体组(t值分别为7.92、4.82、4.72,P<0.01)。分组培养48 h,与单纯高糖组比较,高糖+低浓度外泌体组和高糖+高浓度外泌体组HDF EdU阳性染色率均明显升高(t值分别为5.32、9.88,P<0.01);与高糖+低浓度外泌体组比较,高糖+高浓度外泌体组HDF EdU阳性染色率显著升高(t=5.27,P<0.01)。分组培养48 h,与单纯高糖组比较,高糖+低浓度外泌体组和高糖+高浓度外泌体组HDF中的G0/G1期亚群占比均显著降低(t值分别为3.81、4.31,P<0.05),G2/M+S亚群占比均明显升高(t值分别为3.81、4.31,P<0.05)。分组培养24 h,与单纯高糖组相比,高糖+低浓度外泌体组和高糖+高浓度外泌体组HDF穿过滤膜的数量均明显增多(t值分别为10.14、13.39,P<0.01);与高糖+低浓度外泌体组相比,高糖+高浓度外泌体组HDF穿过滤膜的数量明显增多(t=6.27,P<0.01)。分组培养48 h,与单纯高糖组比较,高糖+低浓度外泌体组和高糖+高浓度外泌体组HDF凋亡率均明显降低(t值分别为3.72、5.53,P<0.05或P<0.01)。分组培养48 h,与单纯高糖组相比,高糖+高浓度外泌体组HDF的miR-145-5p、miR-498 mRNA表达量均明显上升(t值分别为13.03、8.90,P<0.01),miR-503-5p mRNA表达量明显下降(t=3.85,P<0.05);高糖+高浓度外泌体组中HDF的CAMK1D、PTEN基因mRNA表达量均明显低于单纯高糖组(t值分别为8.83、5.97,P<0.01),细胞周期蛋白D1 mRNA表达量明显高于单纯高糖组(t=4.03,P<0.05)。 结论 人dMSC来源外泌体可显著提高高糖老化HDF的增殖和迁移能力,抑制其凋亡。这可能与HDF内miR-145-5p和miR-498表达增高抑制了CAMK1D和PTEN基因的表达及miR-503-5p表达下降促进了细胞周期蛋白D1的表达有关。 Abstract:Objective To establish a high glucose senescent model of human dermal fibroblasts (HDFs), and to investigate the effects of exosomes derived from human decidua mesenchymal stem cells (dMSCs) on the proliferation, migration, and apoptosis of senescent HDFs and possible mechanism. Methods The experimental research method was used. From January to March 2021, discarded foreskin tissue was collected for isolation and culture of primary HDFs from 4 male phimosis patients (aged 18-22 years) admitted for circumcision in the Fourth Medical Center of the PLA General Hospital. The 6th passage of HDFs were taken and divided into low glucose group and high glucose group according to the random number table, and subsequently cultured in low-glucose complete medium and high-glucose complete medium, respectively, with medium changed every 72 h without subculturing. After 10 days of culture, the cells were taken and measured for cellular senescence using the β-galactosidase kit at 24 h after seeding; the expression of senescence-related proteins p16 and p53 was assessed by Western blotting at 48 h after seeding; cell proliferation was detected at 24, 48, and 72 h after seeding using the cell counting kit 8 (CCK-8) method; the cell proliferation was evaluated by 5-ethynyl-2'-deoxyuridine (EdU) staining method, cell cycle and apoptosis were measured by flow cytometry after 48 h of seeding; Transwell experiment was used for the calculation of cell migration rate at 24 h after seeding. The human dMSCs were taken and cultured for 48-72 h from which the exosomes were extracted by differential high speed centrifugal method. The morphology of dMSC exosomes was observed by transmission electron microscopy, the particle size distribution of dMSC exosomes was measured by nanoparticle tracking analysis, and the expression of dMSC-exosomes marker proteins CD9 and tumor susceptibility gene101 (TSG101) were detected by Western blotting. The dMSC exosomes and high-glucose complete medium-induced senescent HDFs were co-cultured for 24 hours, then PKH67 kit was used to detect the uptake of exosomes by HDFs. High-glucose complete medium-induced senescent HDFs were taken and divided into high glucose alone group, high glucose+low concentration of exosomes group, and high glucose+high concentration of exosomes group according to the same method above. The high-glucose complete medium with equal volume of phosphate buffered saline, dMSC exosomes with final concentration of 50 μg/mL, and dMSC exosomes with final concentration of 100 μg/mL were added to the corresponding groups for conventional cell culture, respectively. After grouped, the cell proliferation, cell cycle and apoptosis as well as cell migration were detected by CCK-8 method and EdU staining method, flow cytometry, and Transwell experiment at the corresponding time points as before, respectively. Based on the previous results, high-glucose complete medium-induced senescent HDFs were taken and divided into high glucose alone group and high glucose+high concentration of exosomes group for the same treatment. After being grouped and cultured for 48 h, real-time fluorescent quantitative polymerase chain reaction was used to evaluate the mRNA expression of senescent-related microRNA (miR)-145-5p, miR-498, miR-503-5p, calcium/calmodulin dependent protein kinase 1D (CAMK1D), phosphates and tensin homologue deleted on chromosome ten (PTEN) gene, and Cyclin D1 in high glucose alone group and high glucose+high concentration of exosomes group. Data were statistically analyzed with analysis of variance for factorial design, one-way analysis of variance, least significant difference t test, and independent sample t test. Results At 24 h after seeding, the rate of β-galactosidase-positive staining of HDF in high glucose group was (38.4±4.2)%, which was significantly higher than (16.5±2.2)% of low glucose group (t=4.65, P<0.01). At 48 h after seeding, the expression levels of senescence-related proteins p16 and p53 both were significantly higher in HDFs of high glucose group than those in low glucose group (with t values of 11.85 and 3.02, respectively, P<0.05 or P<0.01). At 0, 24, 48, and 72 h after seeding, the cell proliferation viability of HDFs in high glucose group was all significantly lower than in low glucose group (with t values of 4.13, 9.90, and 15.12, respectively, P<0.01). At 48 h after seeding, the rate of EdU-positive staining of HDFs in high glucose group was obviously lower than that of low glucose group (t=3.83, P<0.05). At 48 h after seeding, the percentage of G2/M+S subpopulations in three subpopulations (G0/G1, S, and G2/M) of HDF cycle was significantly lower in high glucose group than that in low glucose group (t=8.74, P<0.01). At 24 h after seeding, the number of HDFs migrated through the filter membrane to the lower chamber was 37±6 in high glucose group, which was significantly less than 74±7 in low glucose group (t=8.42, P<0.01). At 48 h after seeding, the HDF apoptosis rate was significantly higher in high glucose group than in low glucose group (t=8.48, P<0.01). The dMSC exosomes were cup-shaped or round vesicles with well-defined edges and uniform size distribution. The size of dMSC exosomes was basically in the range of 80-200 nm. Exosomal markers including CD9 and TSG101 were positively presented on the dMSC exosomes. After being co-cultured for 24 hours, the dMSC exosomes were taken up intracellularly by HDFs and mainly distributed around the nucleus of HDFs. After being grouped and cultured for 24, 48, and 72 h, the HDF proliferation viabilities in high glucose+low concentration of exosomes group and high glucose+high concentration of exosomes group were both significantly higher than in high glucose alone group (with t values of 6.36, 6.10, 7.76, 8.92, 12.17, and 10.74, respectively, P<0.01), the HDF proliferation viability in high glucose+high concentration of exosomes group was significantly higher than in high glucose+low concentration of exosomes group (with t values of 7.92, 4.82, and 4.72, respectively, P<0.01). After being grouped and cultured for 48 h, the percentages of EdU-positive HDFs in high glucose+low concentration of exosomes group and high glucose+high concentration of exosomes group were both significantly higher than in high glucose alone group (with t values of 5.32 and 9.88, respectively, P<0.01), the percentage of EdU-positive HDFs in high glucose+high concentration of exosomes group was notably higher than in high glucose+low concentration of exosomes group (t=5.27, P<0.01). After being grouped and cultured for 48 h, the proportion of G0/G1 subpopulation in both high glucose+low concentration of exosomes group and high glucose+high concentration of exosomes group was distinctly lower (with t values of 3.81 and 4.31, respectively, P<0.05), while the proportion of G2/M+S subpopulation was markedly higher (with t values of 3.81, 4.31, respectively, P<0.05) than in high glucose alone group. After being grouped and cultured for 24 h, the number of HDFs migrated through the filter membrane in both high glucose+low concentration of exosomes group and high glucose+high concentration of exosomes group was significantly higher than in high glucose alone group (with t values of 10.14 and 13.39, respectively, P<0.01), the number of HDFs migrated through the filter membrane in high glucose+high concentration of exosomes group was significantly increased than in high glucose+low concentration of exosomes group (t=6.27, P<0.01). After being grouped and cultured for 48 h, the HDF apoptosis rates in high glucose+low concentration of exosomes group and high glucose+high concentration of exosomes group were both significantly lower than in high glucose alone group (with t values of 3.72 and 5.53, respectively, P<0.05 or P<0.01). After being grouped and cultured for 48 h, compared with those in high glucose alone group, the mRNA expression levels of miR-145-5p and miR-498 were both obviously higher (with t values of 13.03 and 8.90, respectively, P<0.01), while the mRNA expression level of miR-503-5p was significantly lower (t=3.85, P<0.05) in high glucose+high concentration of exosomes group. After being grouped and cultured for 48 h, compared with those in high glucose alone group, the mRNA expression levels of CAMK1D and PTEN gene were both significantly lower (with t values of 8.83 and 5.97, respectively, P<0.01), while the mRNA expression level of Cyclin D1 was significantly higher in high glucose+high concentration of exosomes group (t=4.03, P<0.05). Conclusions The dMSC exosomes are capable of improving cell proliferation and migration, and inhibiting cell apoptosis of high-glucose senescent HDFs. This may be related to the mechanism by which the increased expressions of intracellular miR-145-5p and miR-498 inhibit the expression of CAMK1D and PTEN gene, and the decreased expression of miR-503-5p promote the expression of Cyclin D1. -
Key words:
- Mesenchymal stem cells /
- Exosomes /
- Cell aging /
- Glucose solution, hypertonic /
- Fibroblasts
-
皮肤纤维化疾病是一类严重影响患者身心健康的疾病,可由烧创伤、手术或疾病引起 [ 1] ,主要包括增生性瘢痕、瘢痕疙瘩及系统性硬化病等 [ 2, 3, 4] 。皮肤纤维化疾病由于治疗周期长、个体差异大及复发率高等原因,造成了沉重的社会卫生经济负担。目前,临床仍然缺乏能有效缓解或治疗皮肤纤维化疾病的理想药物。
皮肤纤维化疾病的主要病理特点是以胶原蛋白为主的ECM异常沉积,导致瘢痕过度增生或皮肤增厚。Fb在调节ECM稳态中发挥主要作用,其在多条信号通路(包括TGF-β、Wnt和血小板衍生生长因子信号通路)的调控下实现活化和增殖;其次,促炎性细胞因子如TNF-α、IL-1或IL-6等也可作用于Fb促进其活化和增殖 [ 5, 6] 。此外,内皮细胞、巨噬细胞等其他细胞也参与皮肤纤维化疾病的发生与发展 [ 7] 。近年来的相关机制研究为该疾病的治疗带来了新希望,特别是细胞代谢调节领域研究的快速进展,为研究者们提供了新的思考及探索途径。本文就有氧糖酵解相关酶和产物在皮肤纤维化疾病发生与发展中的作用及靶向有氧糖酵解治疗皮肤纤维化疾病的药物进行综述。
1. 皮肤纤维化疾病中异常的有氧糖酵解
糖酵解是指细胞质内葡萄糖在无氧或缺氧条件下分解为乳酸,并产生ATP的过程。在氧气充足的正常细胞中,葡萄糖分解为丙酮酸后易位至线粒体,通过三羧酸循环和氧化呼吸链产生大量的ATP [ 8] 。有氧糖酵解是指即使在氧气充足的情况下,细胞仍然倾向于糖酵解,丙酮酸经乳酸脱氢酶(lactate dehydrogenase,LDH)催化产生大量乳酸,同时抑制线粒体氧化磷酸化(oxidative phosphorylation,OXPHOS),这种现象最初在1927年由Otto Warburg于肿瘤细胞中观察到,并被命名为Warburg效应 [ 9] 。
近年来研究表明,有氧糖酵解与皮肤纤维化疾病的发生与发展密切相关,有氧糖酵解不仅与Fb的增殖、活化相关,也参与ECM的过量积累 [ 10, 11, 12] 。研究显示,在皮肤伤口愈合早期,有氧糖酵解可能作为一种机体自我保护机制,有利于细胞快速供能并促进伤口愈合,但是后期持续增强的有氧糖酵解会影响组织重塑,导致增生性瘢痕形成 [ 13] 。研究者运用氟脱氧葡萄糖正电子发射断层扫描对5例瘢痕疙瘩患者病损处进行扫描,结果显示瘢痕疙瘩较周围正常皮肤葡萄糖聚集增加,提示瘢痕疙瘩中糖代谢发生了改变 [ 14] 。后续研究显示患者瘢痕疙瘩中糖酵解标志物含量明显升高 [ 15] ,瘢痕疙瘩来源的Fb中葡萄糖消耗、乳酸生成、丙酮酸激酶M2(pyruvate kinase M2,PKM2)及Ⅰ型胶原水平均高于正常皮肤Fb [ 16, 17, 18] 。系统性硬化病患者Fb中的糖酵解相关酶储量增加,表明这些细胞在OXPHOS受损的条件下糖酵解水平升高,并产生更多的乳酸 [ 19, 20] 。针对小鼠ECM致密处的足垫皮肤和ECM菲薄的腹部皮肤基因测序结果显示,ECM致密处糖酵解相关分子基因表达水平上调 [ 13] 。这些研究显示,有氧糖酵解在皮肤损伤发生时启动,在整个损伤修复过程中发挥重要作用,但组织重塑期持续增强的有氧糖酵解导致了皮肤纤维化疾病的发生。
2. 有氧糖酵解在皮肤纤维化疾病中的作用
有氧糖酵解在皮肤纤维化疾病中通过促进乳酸生成,调节缺氧诱导因子-1α(hypoxia-inducible factor-1α,HIF-1α)的表达,增强PKM2、葡萄糖转运蛋白1(glucose transporter 1,GLUT1)及磷酸甘油酸激酶1(phosphoglycerate kinase 1,PGK1)的表达,刺激Fb活化和增殖、内皮-间质转化及胶原蛋白合成,从而在皮肤纤维化疾病的发生与发展过程中发挥重要作用。见 图1。
2.1 促进乳酸生成
乳酸由丙酮酸经LDH催化产生,通过单羧酸转运蛋白转运。在纤维化组织中,糖酵解水平的升高导致乳酸大量堆积,促进Fb活化、增殖及内皮-间质转化,增加ECM分泌。有研究表明,人瘢痕疙瘩中央的Fb中LDH、单羧酸转运蛋白4表达水平升高 [ 15] ,这些Fb较正常真皮Fb消耗更多的葡萄糖,产生更多乳酸 [ 21] 。系统性硬化病患者真皮中的Fb有氧糖酵解增强导致乳酸释放增加,随后细胞外酸化促进内皮-间质转化,加重了系统性硬化病中的组织纤维化 [ 20] 。研究者在急性肾损伤小鼠模型中观察到,增强的有氧糖酵解产生的乳酸被肾Fb摄取,诱导Fb活化为肌Fb,引起ECM堆积 [ 22] 。TGF-β 1在Fb转分化为肌Fb中起重要作用,是主要的促纤维化因子之一 [ 23] ,其活化依赖于细胞微环境中的蛋白酶、pH或活性氧 [ 24] 。大量堆积的乳酸可以通过改变人肺Fb微环境pH激活TGF-β 1 [ 25] 。同时,乳酸可加快多种癌细胞系增殖期细胞中有丝分裂的完成,缩短细胞倍增时间,提高增殖效率 [ 26] 。有关肾纤维化的研究表明,乳酸可加快小鼠肾间质Fb的增殖 [ 22] 。进一步的研究揭示了多种癌细胞系和人胚胎肾细胞内乳酸浓度随有丝分裂的进行而增加,乳酸通过在小泛素样修饰物特异性蛋白酶1活性位点与锌形成复合物,结合并抑制小泛素样修饰物特异性蛋白酶1,这一作用稳定了后期促进复合物/细胞周期体亚基4上2个残基的小泛素化修饰,驱动了泛素结合酶E2与后期促进复合物/细胞周期体的结合,刺激细胞周期蛋白B1和分离酶抑制蛋白的定时降解,加快了细胞增殖 [ 26] 。二氯乙酸钠是线粒体丙酮酸脱氢酶激酶(pyruvate dehydrogenase kinase,PDK)的抑制剂,可优先驱动线粒体氧化丙酮酸拮抗乳酸的产生,从而维持细胞周期蛋白B1和分离酶抑制蛋白的稳定,延长有丝分裂的时间,减缓细胞增殖 [ 26] 。
2.2 调节HIF-1α的表达
HIF-1α是调节HIF-1活性的主要亚单位,其可显著提高多种细胞包括Fb的糖酵解水平 [ 27] 。研究者在小鼠胚胎Fb中观察到,HIF-1会上调 PDK1基因表达,抑制丙酮酸脱氢酶,进一步抑制三羧酸循环,使葡萄糖代谢物(丙酮酸)由OXPHOS反应转变为糖酵解 [ 28] 。GLUT对葡萄糖的吸收和转运至关重要,其中GLUT1是分布最为广泛的GLUT。HIF-1作用于多种癌细胞 GLUT1基因的增强子,上调其表达,升高糖酵解水平 [ 29] 。此外,HIF-1α可直接促进Fb活化。关于人肺纤维化的研究表明,肌Fb中6-磷酸果糖-2-激酶/果糖-2,6-二磷酸酶3(6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase 3,PFKFB3)表达增加使得糖酵解水平升高,导致三羧酸循环中间产物琥珀酸水平上调,琥珀酸可稳定HIF-1α的结构,染色质免疫沉淀试验证实HIF-1α在经TGF-β 1处理后直接结合于α平滑肌肌动蛋白基因的启动子区域,促进人肺Fb转分化为肌Fb [ 10] 。
2.3 增强PKM2的表达
PKM2是糖酵解中催化葡萄糖生成丙酮酸的主要限速酶之一。人瘢痕疙瘩Fb中的PKM2在低氧刺激下较在正常Fb中的表达升高 [ 30] 。人增生性瘢痕来源的内源性多肽-增生性瘢痕失调多肽1通过与黏着斑激酶和PKM2结合,下调两者活性,并抑制Smad2磷酸化,达到抑制增生性瘢痕Fb增殖,促进Fb凋亡,降低S期Fb比例及减少胶原合成的作用 [ 31] 。
PKM2以4种不同的酶促状态存在:无活性单体、几乎无活性的二聚体、无活性T态四聚体和活性R态四聚体,四者之间的比例决定PKM2最终的功能 [ 32] 。目前,关于增大PKM2四聚体占比在不同组织纤维化中的作用机制有不同的研究结果。有研究显示,四氯化碳诱导的小鼠肝纤维化模型中肝Fb的PKM2表达上调,敲除 PKM2基因或使PKM2四聚体化会显著抑制体外小鼠肝Fb的活化和增殖 [ 33] 。在选择性PKM2四聚体化激活剂焦磷酸四乙酯-46作用下,小鼠主动脉内皮细胞产生较低浓度的乳酸,抑制了体内外的内皮-间质转化 [ 34] 。然而,PKM2四聚体在人纤维化肺组织中的Fb内表达上调,PKM2四聚体可与Smad7形成络合物,阻断Smad7与TGF-β 1Ⅰ型受体结合,抑制TGF-β 1Ⅰ型受体泛素化,使其保持稳定,从而增强TGF-β 1信号通路转导,促进纤维化进程 [ 35] 。综合以上研究,考虑PKM2在纤维化中发挥不同作用的原因可能基于以下2点:(1)上述研究均未对PKM2四聚体的活性状态进行区分,因此产生了不同的研究结果;(2)PKM2不同构象调控不同的纤维化信号通路,这些信号通路同时受纤维化发展阶段中其他分子的调控。因此,PKM2不同构象在纤维化中的具体作用机制仍需进一步研究明确。
2.4 增强GLUT1的表达
研究显示,烧伤患者皮肤形成的瘢痕疙瘩组织中糖酵解和GLUT1表达增强 [ 36] 。有研究表明,TGF-β 1在小鼠原代肝Fb中可通过经典和非经典通路上调GLUT1的表达,从而促进肝纤维化,通过根皮素抑制GLUT1则可逆转TGF-β 1对肝Fb迁移和增殖的影响,并延迟肝Fb转分化为肌Fb的过程;动物实验结果显示,腹腔注射根皮素缓解了四氯化碳诱导的小鼠肝纤维化 [ 37] 。
2.5 促进PGK1的表达
PGK1在糖酵解过程中催化1,3-二磷酸甘油酸转变成3-磷酸甘油酸,并产生ATP。研究者观察到PGK1在人瘢痕疙瘩组织和瘢痕疙瘩Fb中均高表达,敲除 PGK1基因后,瘢痕疙瘩Fb的增殖、迁移、侵袭和Ⅰ型胶原表达均受到抑制,并且磷脂酰肌醇-3-激酶(phosphoinositide 3-kinase,PI3K)/蛋白激酶B(protein kinase B,Akt)通路的磷酸化被显著抑制,在PI3K抑制剂LY294002作用于瘢痕疙瘩Fb后,GLUT1、LDHA和Ⅰ型胶原的表达均下调,研究者推测PGK1通过PI3K/Akt途径介导上述基因在瘢痕疙瘩Fb中的表达 [ 38] 。
3. 抑制有氧糖酵解的药物
在皮肤纤维化疾病的发生与发展过程中,有氧糖酵解参与并发挥了重要作用,以抑制有氧糖酵解为治疗靶点的药物为皮肤纤维化疾病提供了新的治疗思路。抑制有氧糖酵解的药物通过靶向有氧糖酵解相关酶和综合抑制有氧糖酵解发挥作用。
3.1 靶向有氧糖酵解相关酶的药物
研究显示,紫草素通过抑制PKM2下调糖酵解水平并抑制核苷酸结合寡聚化结构域样受体热蛋白结构域相关蛋白3介导的炎症反应,改善了小鼠烧伤创面的异常愈合,防止了瘢痕疙瘩的形成 [ 36] 。另有研究表明,紫草素可降低机械通气诱导的肺纤维化小鼠肺泡灌洗液中的乳酸和Ⅰ型原胶原羟基端延长肽,减轻肺组织中的胶原沉积,抑制小鼠肺纤维化 [ 39] 。2-脱氧-D-葡萄糖(2-deoxy-D-glucose,2-DG)是己糖激酶-2的抑制剂 [ 11] ,研究者通过构建人真皮原代Fb的Smad3驱动荧光素酶报告基因,使用2-DG抑制糖酵解可下调由TGF-β 1诱导的 Smad3依赖性转录,表明2-DG可下调ECM水平,从而改善皮肤纤维化 [ 13] 。Li等 [ 40] 关于瘢痕疙瘩的研究表明,2-DG可以剂量和时间依赖的方式抑制人瘢痕疙瘩Fb的增殖。3-(3-吡啶基)-1-(4-吡啶基)-2-丙烯-1-酮是PFKFB3的抑制剂 [ 10] ,可缓解由TGF-β 1刺激人真皮Fb发生的胶原沉积 [ 19] 。草氨酸盐可通过竞争性结合LDH而后抑制LDH [ 22] ,继而减少小鼠肾小管上皮细胞的乳酸生成,抑制叶酸诱导损伤后的小鼠肾Fb活化和增殖 [ 22] 。化合物408可抑制LDH5并逆转TGF-β 1介导的人原代肺Fb中代谢方式向有氧糖酵解的转变 [ 41] 。木蝴蝶素A通过抑制LDHA下调人肝Fb内的糖酵解,抑制肝Fb收缩,从而缓解肝纤维化 [ 42] 。
3.2 综合抑制有氧糖酵解的药物
目前研究表明,二甲双胍可下调Warburg效应的关键因子HIF-1α、GLUT1、PDK1、己糖激酶和LDH,在长时程快速心房起搏诱导的犬慢性房颤模型中,通过口服给予二甲双胍,缓解了犬心房纤维化 [ 43] 。另有研究表明,二甲双胍通过调节人肺Fb腺苷酸激活蛋白激酶/哺乳动物雷帕霉素靶蛋白途径抑制人肺Fb胶原合成 [ 44] 。3-溴丙酮酸是一种小分子烷化剂,有研究者在单侧输尿管结扎诱导的小鼠肾纤维化模型中,通过腹腔注射3-溴丙酮酸,使得小鼠肾Fb中有氧糖酵解相关酶己糖激酶-2、LDHA和PKM2的表达水平下调,小鼠肾Fb的活化、增殖和ECM的合成均受到抑制,从而缓解了小鼠肾纤维化 [ 45] 。博来霉素诱导的小鼠肺纤维化研究表明,血管活性肽可通过血管活性肽-MAS相关G蛋白偶联受体D轴下调小鼠肺Fb中己糖激酶-2、PFKFB3表达水平,抑制糖酵解,并缓解博来霉素诱导的小鼠肺纤维化 [ 46] 。雷公藤红素可下调糖酵解酶(如GLUT1、己糖激酶-2、LDHA、PKM2)和相关信号蛋白(如Akt、HIF-1α、哺乳动物雷帕霉素靶蛋白)的表达水平,抑制Warburg效应,减轻小鼠非酒精性脂肪性肝病肝脏纤维化 [ 47] 。
4. 总结与展望
有氧糖酵解在皮肤损伤发生时同步启动,持续存在于整个损伤修复过程中,后期不断增强的有氧糖酵解打破了组织重塑期的能量代谢平衡,通过多种复杂机制促使纤维化发生。使用有氧糖酵解相关酶的抑制剂或其他小分子化合物及目前已经研制出的药物抑制有氧糖酵解,可达到抑制皮肤纤维化的目的,说明有氧糖酵解参与皮肤纤维化的过程并在其中发挥重要作用,抑制有氧糖酵解可以为皮肤纤维化疾病的防治提供一个新方向。但是,尽管目前已有许多关于有氧糖酵解促纤维化相关机制的研究报道,但其大多着眼于内脏器官纤维化,关于皮肤纤维化的研究仍非常有限,缺乏精准的信号通路转导、蛋白相互作用关系及皮肤纤维化疾病发生、发展和稳定过程中的持续监测数据。因此,有氧糖酵解促进皮肤纤维化疾病发生与发展的具体及关键机制的揭秘和整个疾病发展过程中的变化情况亟待探索。相关机制的深入研究会有助于筛选更好的靶向有氧糖酵解的治疗药物,从而为皮肤纤维化疾病提供更精准、更有效的临床治疗方法。
苏建隆:实施研究、采集数据、分析/解释数据、起草文章、统计分析;马奎:对文章的知识性内容作批评性审阅;张翠萍、付小兵:酝酿和设计研究、研究指导、行政支持、技术支持、材料支持所有作者均声明不存在利益冲突巨噬细胞是代谢血红素的主要效应细胞,当受损或衰老红细胞的处理需求增加时,肝脏中巨噬细胞数量会一过性地增加。巨噬细胞在抵御微生物威胁方面也很重要,但血红素过量的病理状态很可能导致免疫抑制的发生。在此,作者揭示了在感染肺炎克雷伯菌后,巨噬细胞需清除的衰老红细胞急剧增多,并最终转变为免疫抑制表型的机制,而免疫抑制表型巨噬细胞的存在,使得肺炎克雷伯菌感染特征呈现为肺外细菌增殖增加,小鼠患脓毒症后的存活率降低。该研究观察到感染缺乏铁载体功能的肺炎克雷伯菌突变体的小鼠与感染野生型菌株的小鼠对过量衰老红细胞的清理结果相似,主要表现为肺细菌负荷没有差异,因此认为肺炎克雷伯菌相关的免疫受损和细菌铁载体获取铁无关。然而,在肺炎克雷伯菌相关脓毒症中,对衰老红细胞的处理导致肝脏转录组组谱呈现出显著的信号转导及转录激活因子1(STAT1)抑制和干扰素相关反应。在感染时,巨噬细胞过度处理血红素致STAT1抑制,这一过程需要核因子E2相关因子1和2的协同激活,但无须血红素加氧酶1诱导。尽管该免疫受损与铁的获取无关,但血红素的卟啉部分足以介导抑制人和小鼠巨噬细胞中STAT1依赖的反应,并促进肺炎克雷伯菌在体内肝脏传播。因此,细胞血红素代谢障碍负向调节STAT1通路与严重感染有关。杨云稀、高晞,编译自《J Clin Invest》, 2021,131(1):e137468;孙炳伟,审校 -
参考文献
(40) [1] LiB, BianX, HuW, et al. Regenerative and protective effects of calcium silicate on senescent fibroblasts induced by high glucose[J]. Wound Repair Regen,2020,28(3):315-325. DOI: 10.1111/wrr.12794. [2] LiM,ZhaoY,HaoH,et al.Umbilical cord-derived mesenchymal stromal cell-conditioned medium exerts in vitro antiaging effects in human fibroblasts[J].Cytotherapy,2017,19(3):371-383.DOI: 10.1016/j.jcyt.2016.12.001. [3] RaniS,RyanAE,GriffinMD,et al.Mesenchymal stem cell-derived extracellular vesicles: toward cell-free therapeutic applications[J].Mol Ther,2015,23(5):812-823.DOI: 10.1038/mt.2015.44. [4] NikfarjamS,RezaieJ,ZolbaninNM,et al.Mesenchymal stem cell derived-exo somes: a modern approach in translational medicine[J].J Transl Med,2020,18(1):449.DOI: 10.1186/s12967-020-02622-3. [5] KomakiM,NumataY,MoriokaC,et al.Exosomes of human placenta-derived mesenchymal stem cells stimulate angiogenesis[J].Stem Cell Res Ther,2017,8(1):219.DOI: 10.1186/s13287-017-0660-9. [6] ZhaoB,ZhangY,HanS,et al.Exosomes derived from human amniotic epithelial cells accelerate wound healing and inhibit scar formation[J].J Mol Histol,2017,48(2):121-132.DOI: 10.1007/s10735-017-9711-x. [7] DashBC,XuZ,LinL,et al.Stem cells and engineered scaffolds for regenerative wound healing[J].Bioengineering (Basel),2018, 5(1):23. DOI: 10.3390/bioengineering5010023. [8] LermanOZ,GalianoRD,ArmourM,et al.Cellular dysfunction in the diabetic fibroblast: impairment in migration, vascular endothelial growth factor production, and response to hypoxia[J].Am J Pathol,2003,162(1):303-312.DOI: 10.1016/S0002-9440(10)63821-7. [9] ThomasK,KiwitM,KernerW.Glucose concentration in human subcutaneous adipose tissue: comparison between forearm and abdomen[J].Exp Clin Endocrinol Diabetes,1998,106(6):465-469.DOI: 10.1055/s-0029-1212017. [10] WeiQ,WangY,MaK,et al.Extracellular vesicles from human umbilical cord mesenchymal stem cells facilitate diabetic wound healing through miR-17-5p-mediated enhancement of angiogenesis[J/OL].Stem Cell Rev Rep,2021(2021-05-04)[2021-10-13]. https://pubmed.ncbi.nlm.nih.gov/33942217/. DOI:10.1007/s12015-021-10176-0. [published online ahead of print]. [11] ChengL,ZhangK,WuS,et al.Focus on mesenchymal stem cell-derived exosomes: opportunities and challenges in cell-free therapy[J].Stem Cells Int,2017,2017:6305295.DOI: 10.1155/2017/6305295. [12] LuK,LiHY,YangK,et al.Exosomes as potential alternatives to stem cell therapy for intervertebral disc degeneration: in-vitro study on exosomes in interaction of nucleus pulposus cells and bone marrow mesenchymal stem cells[J].Stem Cell Res Ther,2017,8(1):108.DOI: 10.1186/s13287-017-0563-9. [13] Casado-DíazA,Quesada-GómezJM,DoradoG.Extracellular vesicles derived from mesenchymal stem cells (MSC) in regenerative medicine: applications in skin wound healing[J].Front Bioeng Biotechnol,2020,8:146.DOI: 10.3389/fbioe.2020.00146. [14] HuY,RaoSS,WangZX,et al.Exosomes from human umbilical cord blood accelerate cutaneous wound healing through miR-21-3p-mediated promotion of angiogenesis and fibroblast function[J].Theranostics,2018,8(1):169-184.DOI: 10.7150/thno.21234. [15] PomattoM,GaiC,NegroF,et al.Differential therapeutic effect of extracellular vesicles derived by bone marrow and adipose mesenchymal stem cells on wound healing of diabetic ulcers and correlation to their cargoes[J].Int J Mol Sci,2021, 22(8):3851. DOI: 10.3390/ijms22083851. [16] WangC,WangM,XuT,et al.Engineering bioactive self-healing antibacterial exosomes hydrogel for promoting chronic diabetic wound healing and complete skin regeneration[J].Theranostics,2019,9(1):65-76.DOI: 10.7150/thno.29766. [17] McBrideJD,Rodriguez-MenocalL,GuzmanW,et al.Bone marrow mesenchymal stem cell-derived CD63+ exosomes transport Wnt3a exteriorly and enhance dermal fibroblast proliferation, migration, and angiogenesis in vitro[J].Stem Cells Dev,2017,26(19):1384-1398.DOI: 10.1089/scd.2017.0087. [18] YangJ,ChenZ,PanD,et al.Umbilical cord-derived mesenchymal stem cell-derived exosomes combined pluronic F127 hydrogel promote chronic diabetic wound healing and complete skin regeneration[J].Int J Nanomedicine,2020,15:5911-5926.DOI: 10.2147/IJN.S249129. [19] ZhangW,BaiX,ZhaoB,et al.Cell-free therapy based on adipose tissue stem cell-derived exosomes promotes wound healing via the PI3K/Akt signaling pathway[J].Exp Cell Res,2018,370(2):333-342.DOI: 10.1016/j.yexcr.2018.06.035. [20] FangS,XuC,ZhangY,et al.Umbilical cord-derived mesenchymal stem cell-derived exosomal microRNAs suppress myofibroblast differentiation by inhibiting the transforming growth factor-β/SMAD2 pathway during wound healing[J].Stem Cells Transl Med,2016,5(10):1425-1439.DOI: 10.5966/sctm.2015-0367. [21] HuL,WangJ,ZhouX,et al.Exosomes derived from human adipose mensenchymal stem cells accelerates cutaneous wound healing via optimizing the characteristics of fibroblasts[J].Sci Rep,2016,6:32993.DOI: 10.1038/srep32993. [22] WiklanderOP, NordinJZ, O'LoughlinA,et al.Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting[J].J Extracell Vesicles,2015,4:26316.DOI: 10.3402/jev.v4.26316. [23] YangD,ZhangW,ZhangH,et al.Progress, opportunity, and perspective on exosome isolation - efforts for efficient exosome-based theranostics[J].Theranostics,2020,10(8):3684-3707.DOI: 10.7150/thno.41580. [24] DingM,WangC,LuX,et al.Comparison of commercial exosome isolation kits for circulating exosomal microRNA profiling[J].Anal Bioanal Chem,2018,410(16):3805-3814.DOI: 10.1007/s00216-018-1052-4. [25] KusumaGD,CarthewJ,LimR,et al.Effect of the microenvironment on mesenchymal stem cell paracrine signaling: opportunities to engineer the therapeutic effect[J].Stem Cells Dev,2017,26(9):617-631.DOI: 10.1089/scd.2016.0349. [26] WuD,KangL,TianJ,et al.Exosomes derived from bone mesenchymal stem cells with the stimulation of Fe3O4 nanoparticles and static magnetic field enhance wound healing through upregulated miR-21-5p[J].Int J Nanomedicine,2020,15:7979-7993.DOI: 10.2147/IJN.S275650. [27] YangH,LinJ,JiangJ,et al.miR-20b-5p functions as tumor suppressor microRNA by targeting cyclinD1 in colon cancer[J].Cell Cycle,2020,19(21):2939-2954.DOI: 10.1080/15384101.2020.1829824. [28] DimitrovaN,GochevaV,BhutkarA,et al.Stromal expression of miR-143/145 promotes neoangiogenesis in lung cancer development[J].Cancer Discov,2016,6(2):188-201.DOI: 10.1158/2159-8290.CD-15-0854. [29] DimitrovaN,GochevaV,BhutkarA,et al.Stromal expression of miR-143/145 promotes neoangiogenesis in lung cancer development[J].Cancer Discov,2016,6(2):188-201.DOI: 10.1158/2159-8290.CD-15-0854. [30] LawsonJ,DickmanC,MacLellanS,et al.Selective secretion of microRNAs from lung cancer cells via extracellular vesicles promotes CAMK1D-mediated tube formation in endothelial cells[J].Oncotarget,2017,8(48):83913-83924.DOI: 10.18632/oncotarget.19996. [31] GrasC,RatusznyD,HadamitzkyC,et al.miR-145 contributes to hypertrophic scarring of the skin by inducing myofibroblast activity[J].Mol Med,2015,21(1):296-304.DOI: 10.2119/molmed.2014.00172. [32] ChaiC,WuH,WangB,et al.MicroRNA-498 promotes proliferation and migration by targeting the tumor suppressor PTEN in breast cancer cells[J].Carcinogenesis,2018,39(9):1185-1196.DOI: 10.1093/carcin/bgy092. [33] DuanXM,LiuXN,LiYX,et al.MicroRNA-498 promotes proliferation, migration, and invasion of prostate cancer cells and decreases radiation sensitivity by targeting PTEN[J].Kaohsiung J Med Sci,2019,35(11):659-671.DOI: 10.1002/kjm2.12108. [34] LiJ,ZhangF,LiH,et al.Circ_0010220-mediated miR-503-5p/CDCA4 axis contributes to osteosarcoma progression tumorigenesis[J].Gene,2020,763:145068.DOI: 10.1016/j.gene.2020.145068. [35] MarkopoulosGS,RoupakiaE,TokamaniM,et al.Senescence-associated microRNAs target cell cycle regulatory genes in normal human lung fibroblasts[J].Exp Gerontol,2017,96:110-122.DOI: 10.1016/j.exger.2017.06.017. [36] JiangL,ZhaoZ,ZhengL,et al.Downregulation of miR-503 promotes ESCC cell proliferation, migration, and invasion by targeting cyclin D1[J].Genomics Proteomics Bioinformatics,2017,15(3):208-217.DOI: 10.1016/j.gpb.2017.04.003. [37] BovyN,BlommeB,FrèresP,et al.Endothelial exosomes contribute to the antitumor response during breast cancer neoadjuvant chemotherapy via microRNA transfer[J].Oncotarget,2015,6(12):10253-10266.DOI: 10.18632/oncotarget.3520. [38] HouSQ,OuyangM,BrandmaierA,et al.PTEN in the maintenance of genome integrity: From DNA replication to chromosome segregation[J].Bioessays,2017,39(10):10. DOI: 10.1002/bies.201700082. [39] LiB,LuanS,ChenJ,et al.The MSC-derived exosomal incRNA H19 promotes wound healing in diabetic foot ulcers by upregulating PTEN via microRNA-152-3p[J].Mol Ther Nucleic Acids,2020,19:814-826.DOI: 10.1016/j.omtn.2019.11.034. [40] LongJ,OuC,XiaH,et al.MiR-503 inhibited cell proliferation of human breast cancer cells by suppressing CCND1 expression[J].Tumour Biol,2015,36(11):8697-8702.DOI: 10.1007/s13277-015-3623-8. -
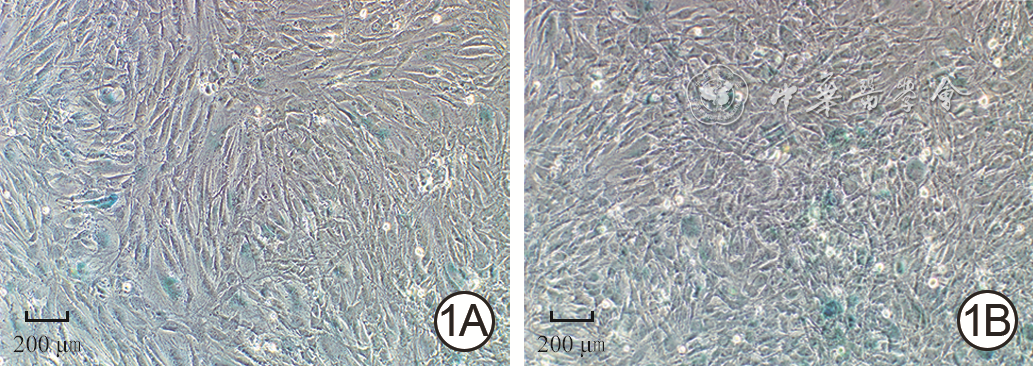
1 2组人真皮成纤维细胞中接种后24 h β-半乳糖苷酶表达情况 光学显微镜×100,图中标尺为200 μm。1A.低糖组β-半乳糖苷酶染色;1B.高糖组β-半乳糖苷酶阳性染色率较图1A高
注:低糖组、高糖组细胞培养基中的葡萄糖物质的量浓度分别为5.5、35.0 mmol/L;图中蓝绿色指示细胞β-半乳糖苷酶阳性染色
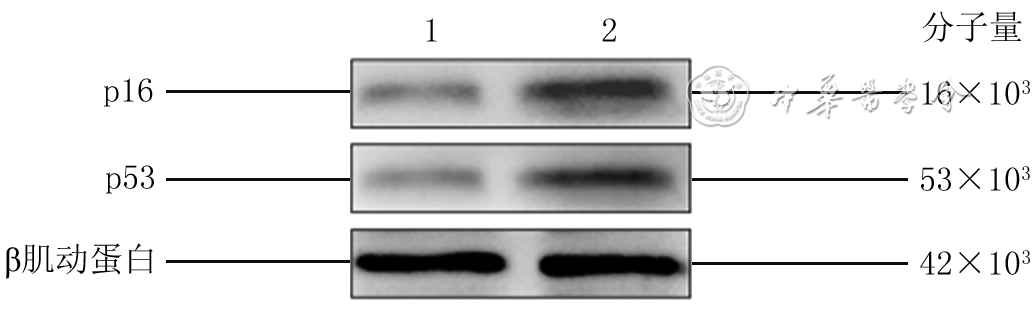
2 蛋白质印迹法检测2组人真皮成纤维细胞接种后48 h衰老相关蛋白表达
注:1为低糖组,2为高糖组;低糖组、高糖组细胞培养基中的葡萄糖物质的量浓度分别为5.5、35.0 mmol/L
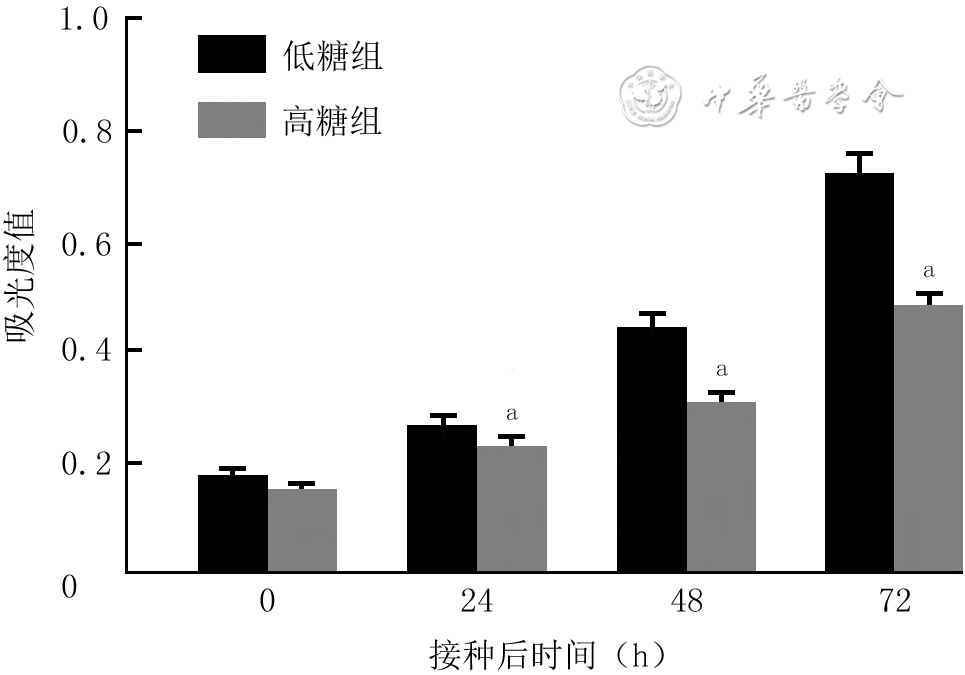
3 细胞计数试剂盒8法检测2组人真皮成纤维细胞接种各时间点的增殖活性(样本数为3,
注:低糖组、高糖组细胞培养基中的葡萄糖物质的量浓度分别为5.5、35.0 mmol/L;处理因素主效应,F=244.90,P<0.001;时间因素主效应,F=892.30,P<0.001;两者交互作用,F=56.23,P<0.001;与低糖组比较,aP<0.01
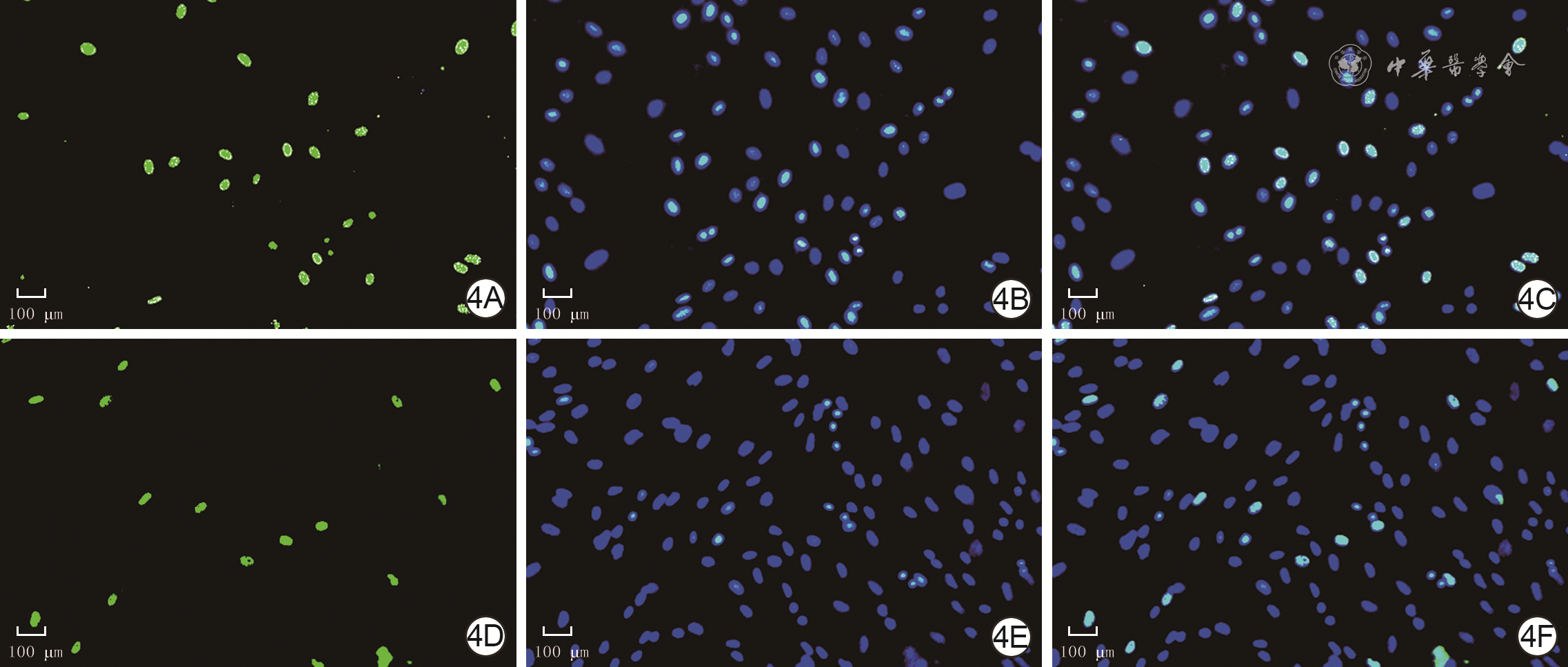
4 EdU染色法检测2组人真皮成纤维细胞培养48 h增殖活力 EdU-hochst×200,图中标尺为100 μm。4A、4B、4C.分别为低糖组细胞EdU染色、细胞核染色、细胞EdU与细胞核共染色情况;4D、4E、4F.分别为高糖组细胞EdU染色、细胞核染色、细胞EdU与细胞核共染色情况,细胞核完整,图4D EdU阳性细胞较图4A明显减少
注:低糖组、高糖组细胞培养基中的葡萄糖浓度分别为5.5、35.0 mmol/L;以细胞脱氧尿嘧啶核苷(EdU)阳性染色为绿色,细胞核阳性染色为蓝色,绿色+蓝色双荧光染色为增殖细胞
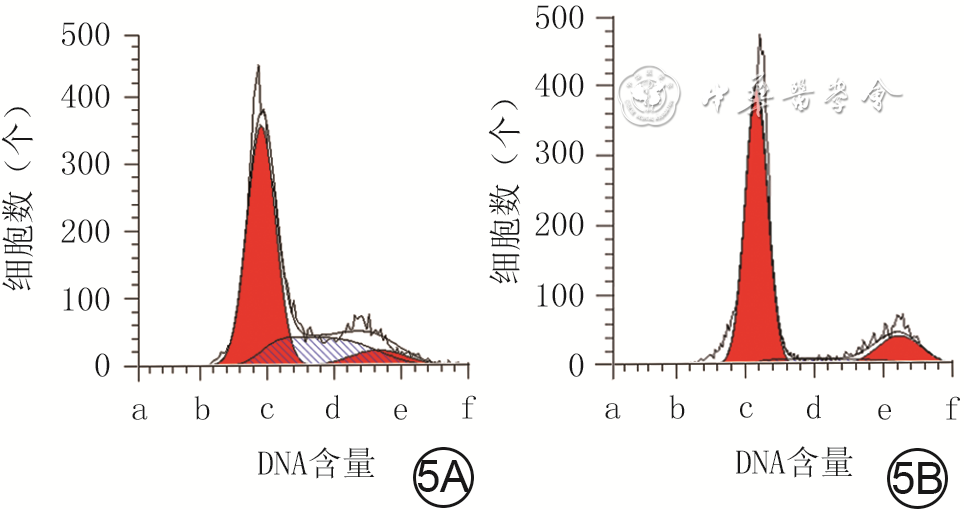
5 流式细胞术检测2组人真皮成纤维细胞培养48 h的细胞周期情况。5A、5B.分别为低糖组、高糖组
注:低糖组、高糖组细胞培养基中的葡萄糖物质的量浓度分别为5.5、35.0 mmol/L;图5A、5B中横坐标a、b、c、d、e、f分别为0、500、1 000、1 600、2 000、2 500;图中第1个红色波峰为G1期,第2个红色波峰为G2期,蓝色斜线区为S期
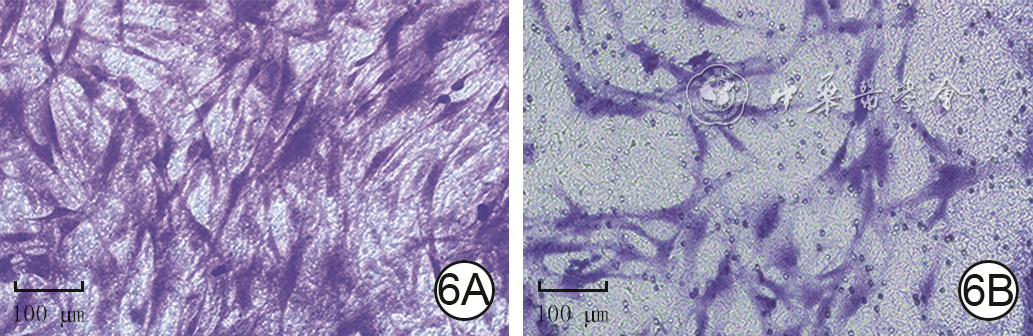
6 Transwell实验观察2组人真皮成纤维细胞培养24 h迁移情况 结晶紫×200,图中标尺为100 μm。6A、6B.分别为低糖组、高糖组,图6B穿膜细胞数较图6A少
注:低糖组、高糖组细胞培养基中的葡萄糖物质的量浓度分别为5.5、35.0 mmol/L;图中紫色指示穿膜细胞
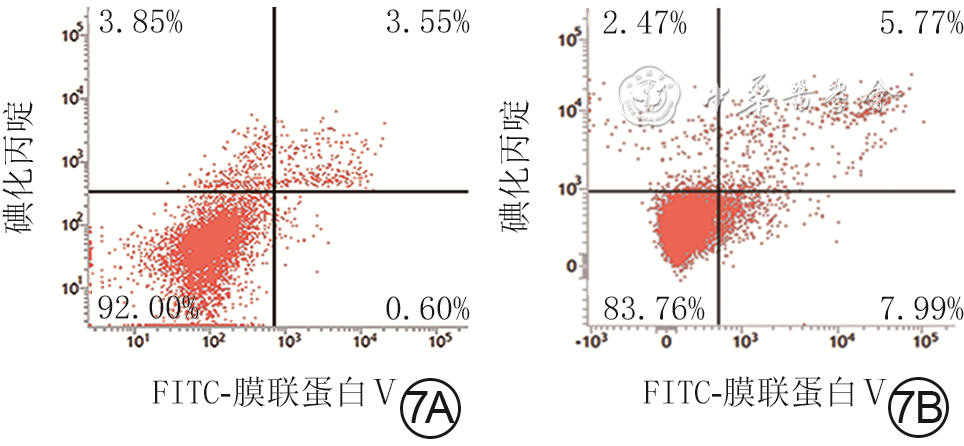
7 流式细胞术检测2组人真皮成纤维细胞培养48 h凋亡情况。7A.低糖组绝大多数细胞处于左下象限,凋亡细胞少;7B.高糖组细胞右上、右下象限凋亡细胞数较图7A明显增多
注:低糖组、高糖组细胞培养基中的葡萄糖物质的量浓度分别为5.5、35.0 mmol/L;FITC为异硫氰酸荧光素;图中左下象限显示活细胞,右下象限显示早期凋亡细胞,右上象限显示晚期凋亡细胞和坏死细胞,左上象限显示细胞收集过程中出现的损伤细胞
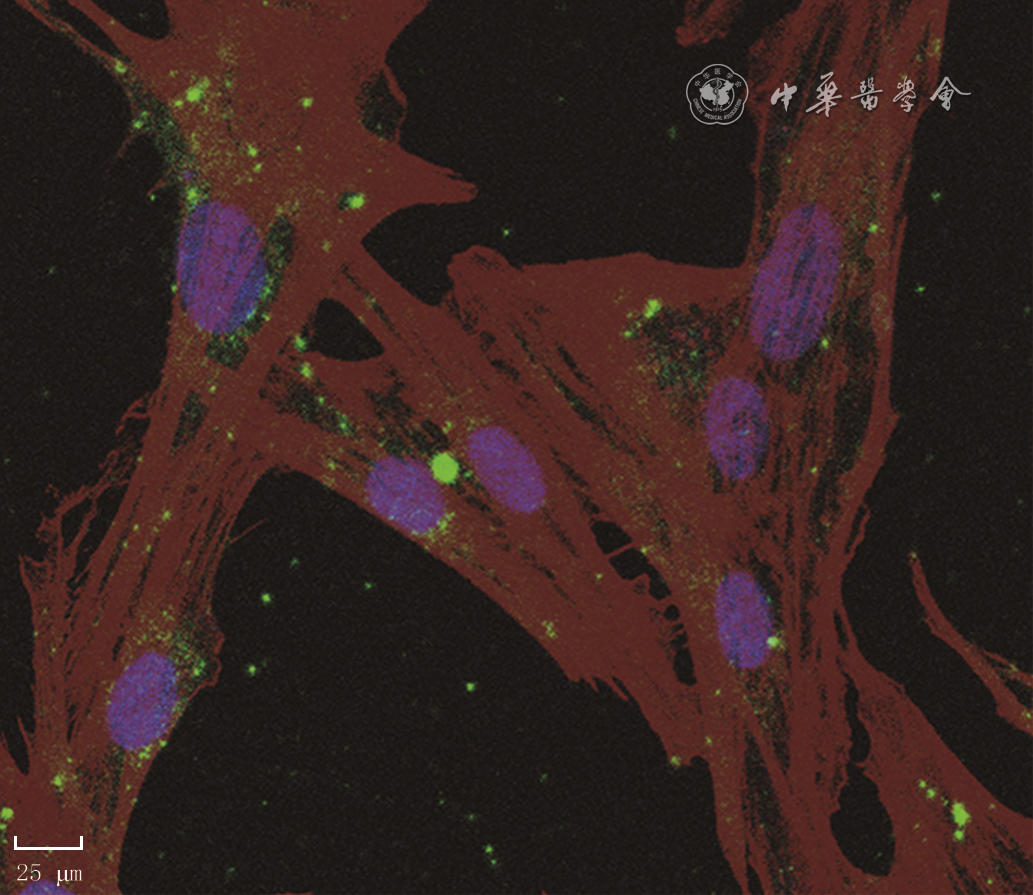
11 人真皮成纤维细胞(HDF)与人dMSC外泌体共培养24 h后,外泌体被HDF内吞入细胞质,并聚集于细胞核周围培养 4',6-二脒基-2-苯基吲哚-罗丹明标记的鬼笔环肽-PKH67×400,图中标尺为25 μm
注:紫色指示细胞核,酱红色指示细胞骨架,绿色为人蜕膜间充质干细胞(dMSC)来源外泌体

12 细胞计数试剂盒8法检测3组人真皮成纤维细胞培养各时间点活力(样本数为3,
注:单纯高糖组、高糖+低浓度外泌体组、高糖+高浓度外泌体组分别于高糖完全培养基中加入等体积的磷酸盐缓冲液、终质量浓度50 μg/mL人蜕膜间充质干细胞外泌体、终质量浓度100 μg/mL 人蜕膜间充质干细胞外泌体进行常规细胞培养;处理因素主效应,F=107.80,P<0.001;时间因素主效应,F=2 992.00,P<0.001;两者交互作用,F=24.99,P<0.001;与单纯高糖组比较,aP<0.01;与高糖+低浓度外泌体组比较,bP<0.01

13 EdU染色法检测3组HDF培养48 h的增殖活力 EdU-hoechst×200,图中标尺为100 μm。13A、13B、13C.分别为单纯高糖组、高糖+低浓度外泌体组、高糖+高浓度外泌体组细胞EdU与细胞核共染色情况,细胞核完整,图13B EdU阳性细胞较图13A明显增多,且图13C EdU阳性染色细胞数最多
注:单纯高糖组、高糖+低浓度外泌体组、高糖+高浓度外泌体组分别于高糖完全培养基中加入等体积的磷酸盐缓冲液、终质量浓度50 μg/mL人蜕膜间充质干细胞外泌体、终质量浓度100 μg/mL人蜕膜间充质干细胞外泌体进行常规细胞培养;细胞脱氧尿嘧啶核苷(EdU)阳性染色为绿色,细胞核阳性染色为蓝色,绿色+蓝色双荧光染色为增殖的人真皮成纤维细胞(HDF)
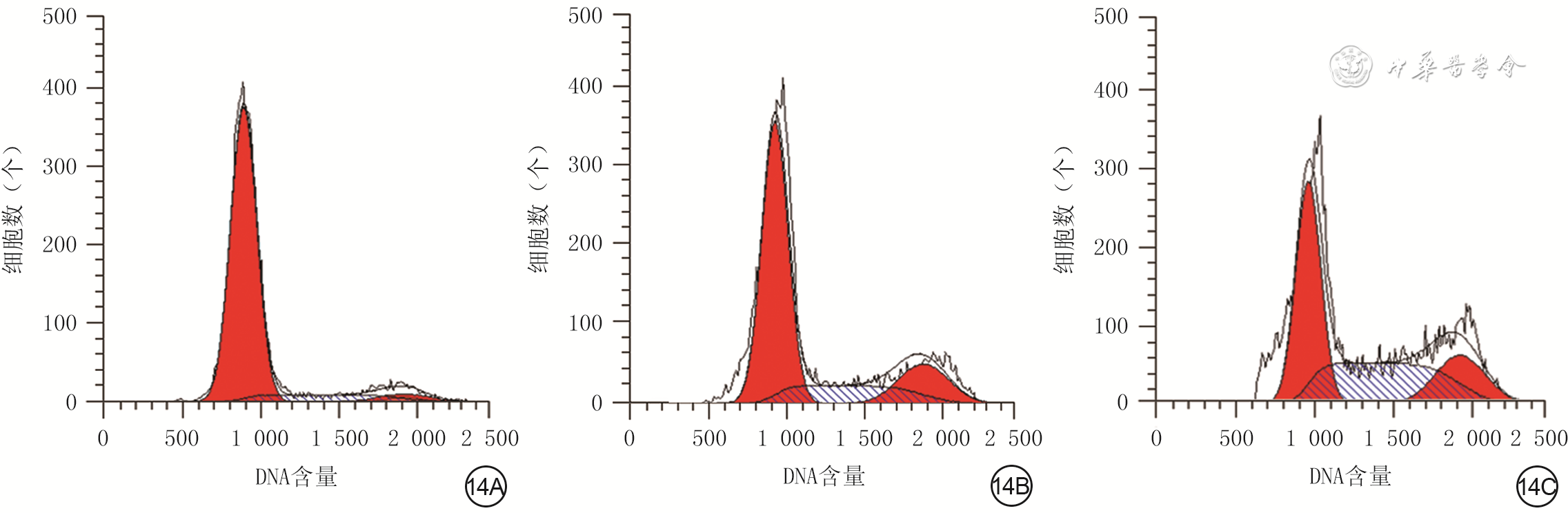
14 流式细胞术检测3组人真皮成纤维细胞培养48 h的细胞周期情况。14A、14B、14C.分别为单纯高糖组、高糖+低浓度外泌体组、高糖+高浓度外泌体组
注:单纯高糖组、高糖+低浓度外泌体组、高糖+高浓度外泌体组分别于高糖完全培养基中加入等体积的磷酸盐缓冲液、终质量浓度50 μg/mL人蜕膜间充质干细胞外泌体、终质量浓度100 μg/mL人蜕膜间充质干细胞外泌体进行常规细胞培养;图中第1个红色波峰为G1期,第2个红色波峰为G2期,蓝色斜线区为S期

15 Transwell实验观察3组人真皮成纤维细胞培养24 h迁移能力 结晶紫×200,图中标尺为100 μm。15A、15B、15C.分别为单纯高糖组、高糖+低浓度外泌体组、高糖+高浓度外泌体组穿膜细胞生长情况,图15B、15C穿膜细胞数均较图15A多,且图15C穿膜细胞数最多
注:单纯高糖组、高糖+低浓度外泌体组、高糖+高浓度外泌体组分别于高糖完全培养基中加入等体积的磷酸盐缓冲液、终质量浓度50 μg/mL人蜕膜间充质干细胞外泌体、终质量浓度100 μg/mL人蜕膜间充质干细胞外泌体进行常规细胞培养;图中紫色指示穿膜细胞
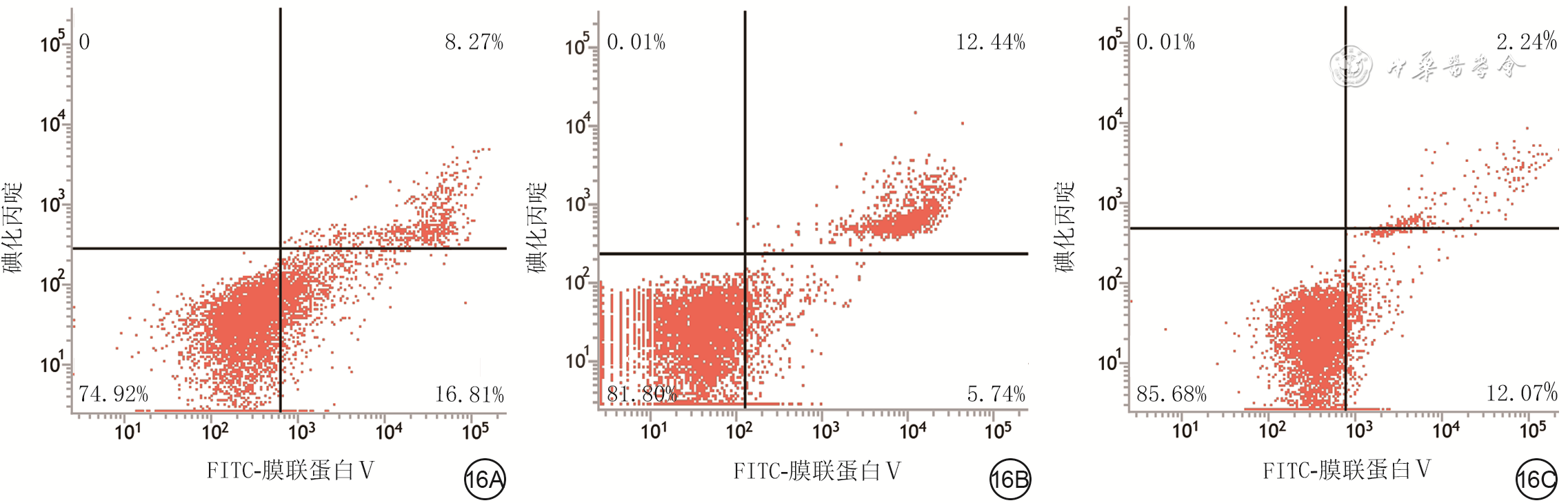
16 流式细胞术检测3组人真皮成纤维细胞(HDF)培养48 h凋亡情况。16A.单纯高糖组HDF绝大多数细胞处于左下象限,凋亡细胞较多;16B.高糖+低浓度外泌体组凋亡HDF较单纯高糖组减少;16C.高糖+高浓度外泌体组右上/下象限凋亡HDF较图16A明显减少,且凋亡细胞数低于图16B
注:单纯高糖组、高糖+低浓度外泌体组、高糖+高浓度外泌体组分别于高糖完全培养基中加入等体积的磷酸盐缓冲液、终质量浓度50 μg/mL人蜕膜间充质干细胞外泌体、终质量浓度100 μg/mL人蜕膜间充质干细胞外泌体进行常规细胞培养;FITC为异硫氰酸荧光素;图中左下象限显示活细胞,右下象限显示早期凋亡细胞,右上象限显示晚期凋亡细胞和坏死细胞,左上象限显示细胞收集过程中出现的损伤细胞
表1 2组高糖诱导老化人真皮成纤维细胞培养48 h衰老相关微小RNA表达情况比较(
组别 样本数 miR-145-5p miR-498 miR-503-5p 单纯高糖组 3 1.01±0.14 1.06±0.21 1.05±0.37 高糖+高浓度外泌体组 3 12.15±1.47 2.89±0.29 0.22±0.03 t值 13.03 8.90 3.85 P值 <0.001 <0.001 0.018 注:单纯高糖组、高糖+高浓度外泌体组分别于高糖完全培养基中加入等体积的磷酸盐缓冲液、终质量浓度100 μg/mL人蜕膜间充质干细胞外泌体进行常规细胞培养;miR为微小RNA  下载: 导出CSV
下载: 导出CSV
表2 2组高糖诱导老化人真皮成纤维细胞中衰老相关基因的mRNA表达情况比较(
组别 样本数 CAMK1D PTEN基因 细胞周期蛋白D1 单纯高糖组 3 1.00±0.11 1.00±0.04 1.14±0.18 高糖+高浓度外泌体组 3 0.37±0.06 0.52±0.13 2.85±0.70 t值 8.83 5.97 4.03 P值 <0.001 0.003 <0.015 注:单纯高糖组、高糖+高浓度外泌体组分别于高糖完全培养基中加入等体积的磷酸盐缓冲液、终质量浓度100 μg/mL人蜕膜间充质干细胞外泌体进行常规细胞培养;CAMK1D为钙/钙调素依赖性蛋白激酶1D,PTEN基因为人第10号染色体缺失的磷酸酶及张力蛋白同源的基因
下载: 导出CSV
-


 下载:
下载:


计量
- 文章访问数: 521
- HTML全文浏览量: 153
- PDF下载量: 34
- 被引次数: 0
