



Regulatory effects of bio-intensity electric field on microtubule acetylation in human epidermal cell line HaCaT
-
摘要:
目的 探讨生物强度电场对人表皮细胞株HaCaT方向性迁移及微管乙酰化水平的调节作用,以期为临床创面修复提供分子理论依据。 方法 采用实验研究方法。取HaCaT细胞,分为置于电场装置中不通电处理3 h的模拟电场组和用强度200 mV/mm电场处理3 h的电场处理组(处理方法下同,样本数分别为54、52),在活细胞工作站中观察处理3 h内细胞运动的方向并计算细胞运动的位移速度、轨迹速度及运动方向性cosθ。取2批HaCaT细胞,分为模拟电场组和用强度200 mV/mm的电场处理相应时间的电场处理1 h组、电场处理2 h组、电场处理3 h组及模拟电场组和用相应强度的电场处理3 h的100 mV/mm电场组、200 mV/mm电场组、300 mV/mm电场组,采用蛋白质印迹法检测乙酰化α-微管蛋白的表达(样本数均为3)。取HaCaT细胞,分为模拟电场组和电场处理组,采用免疫荧光法观测乙酰化α-微管蛋白的表达及定位(样本数为3)。对数据行Kruskal-Wallis H检验、Mann-Whitney U检验、Bonferroni校正、单因素方差分析、LSD检验及独立样本t检验。 结果 处理3 h内,与模拟电场组相比,电场处理组细胞有明显定向迁移的趋势,位移速度、轨迹速度均明显加快(Z值分别为-8.53、-2.05,P<0.05或P<0.01),方向性明显增强(Z=-8.65,P<0.01)。与模拟电场组(0.80±0.14)比较,电场处理1 h组、电场处理2 h组细胞中乙酰化α-微管蛋白表达量(1.50±0.08、1.89±0.06)均无明显变化(P>0.05),电场处理3 h组细胞乙酰化α-微管蛋白表达量(3.37±0.36)明显增高(Z=-3.06,P<0.05)。处理3 h,100 mV/mm电场组、200 mV/mm电场组、300 mV/mm电场组细胞乙酰化α-微管蛋白表达量分别为1.63±0.05、2.24±0.08、2.00±0.13,均较模拟电场组的0.95±0.27明显增高(P<0.01);200 mV/mm电场组、300 mV/mm电场组细胞乙酰化α-微管蛋白表达量均较100 mV/mm电场组明显增高(P<0.01);300 mV/mm电场组细胞乙酰化α-微管蛋白表达量较200 mV/mm电场组明显降低(P<0.05)。处理3 h,电场处理组细胞中乙酰化α-微管蛋白的分布较模拟电场组更具有方向性,电场处理组细胞乙酰化α-微管蛋白的表达量较模拟电场组明显增高(t=5.78,P<0.01)。 结论 生物强度电场可促进HaCaT细胞定向迁移,在200 mV/mm电场强度下处理3 h可明显促进微管乙酰化水平。 Abstract:Objective To investigate the regulatory effects of bio-intensity electric field on directional migration and microtubule acetylation in human epidermal cell line HaCaT, aiming to provide molecular theoretical basis for the clinical treatment of wound repair. Methods The experimental research methods were used. HaCaT cells were collected and divided into simulated electric field group (n=54) placed in the electric field device without electricity for 3 h and electric field treatment group (n=52) treated with 200 mV/mm electric field for 3 h (the same treatment methods below). The cell movement direction was observed in the living cell workstation and the movement velocity, trajectory velocity, and direction of cosθ of cell movement within 3 h of treatment were calculated. HaCaT cells were divided into simulated electric field group and electric field treatment 1 h group, electric field treatment 2 h group, and electric field treatment 3 h group which were treated with 200 mV/mm electric field for corresponding time. HaCaT cells were divided into simulated electric field group and 100 mV/mm electric field group, 200 mV/mm electric field group, and 300 mV/mm electric field group treated with electric field of corresponding intensities for 3 h. The protein expression of acetylated α-tubulin was detected by Western blotting (n=3). HaCaT cells were divided into simulated electric field group and electric field treatment group, and the protein expression of acetylated α-tubulin was detected and located by immunofluorescence method (n=3). Data were statistically analyzed with Kruskal-Wallis H test,Mann-Whitney U test, Bonferroni correction, one-way analysis of variance, least significant difference test, and independent sample t test. Results Within 3 h of treatment, compared with that in simulated electric field group, the cells in electric field treatment group had obvious tendency to move directionally, the movement velocity and trajectory velocity were increased significantly (with Z values of -8.53 and -2.05, respectively, P<0.05 or P<0.01), and the directionality was significantly enhanced (Z=-8.65, P<0.01). Compared with (0.80±0.14) in simulated electric field group, the protein expressions of acetylated α-tubulin in electric field treatment 1 h group (1.50±0.08) and electric field treatment 2 h group (1.89±0.06) were not changed obviously (P>0.05), while the protein expression of acetylated α-tubulin of cells in electric field treatment 3 h group (3.37±0.36) was increased significantly (Z=-3.06, P<0.05). After treatment for 3 h, the protein expressions of acetylated α-tubulin of cells in 100 mV/mm electric field group, 200 mV/mm electric field group, and 300 mV/mm electric field group were 1.63±0.05, 2.24±0.08, and 2.00±0.13, respectively, which were significantly more than 0.95±0.27 in simulated electric field group (P<0.01). Compared with that in 100 mV/mm electric field group, the protein expressions of acetylated α-tubulin in 200 mV/mm electric field group and 300 mV/mm electric field group were increased significantly (P<0.01); the protein expression of acetylated α-tubulin of cells in 300 mV/mm electric field group was significantly lower than that in 200 mV/mm electric field group (P<0.05). After treatment for 3 h, compared with that in simulated electric field group, the acetylated α-tubulin of cells had enhanced directional distribution and higher protein expression (t=5.78, P<0.01). Conclusions Bio-intensity electric field can induce the directional migration of HaCaT cells and obviously up-regulate the level of α-ubulin acetylation after treatment at 200 mV/mm bio-intensity electric field for 3 h. -
Key words:
- Skin /
- Cell movement /
- Tubulin /
- Acetylation /
- Bio-intensity electric field /
- Epidermal cells
-
参考文献
(37) [1] GradaA, Otero-VinasM, Prieto-CastrilloF, et al. Research techniques made simple: analysis of collective cell migration using the wound healing assay[J]. J Invest Dermatol, 2017, 137(2): e11-e16. DOI: 10.1016/j.jid.2016.11.020. [2] 冀然,张泽,王文平,等.生物强度电场对人表皮细胞株 HaCaT和小鼠表皮细胞运动性及CD9表达的影响[J].中华烧伤杂志,2021,37(1):34-41.DOI: 10.3760/cma.j.cn501120-20200115-00023. [3] TaiG, TaiM, ZhaoM. Electrically stimulated cell migration and its contribution to wound healing[J/OL]. Burns Trauma, 2018, 6(1): 20[2022-10-23]. https://pubmed.ncbi.nlm.nih.gov/30003115/. DOI: 10.1186/s41038-018-0123-2. [4] 王文平,冀然,张泽,等.生物强度电场对人皮肤成纤维细胞转化的调节作用[J].中华烧伤与创面修复杂志,2022,38(4):354-362.DOI: 10.3760/cma.j.cn501120-20210112-00017. [5] JiR,TengM,ZhangZ,et al.Electric field down-regulates CD9 to promote keratinocytes migration through AMPK pathway[J].Int J Med Sci,2020,17(7):865-873.DOI: 10.7150/ijms.42840. [6] LinBJ,TsaoSH,ChenA,et al.Lipid rafts sense and direct electric field-induced migration[J].Proc Natl Acad Sci U S A,2017,114(32):8568-8573.DOI: 10.1073/pnas.1702526114. [7] GarcinC,StraubeA.Microtubules in cell migration[J].Essays Biochem,2019,63(5):509-520.DOI: 10.1042/EBC20190016. [8] JankeC, MontagnacG. Causes and consequences of microtubule acetylation[J]. Curr Biol, 2017, 27(23):R1287-R1292. DOI: 10.1016/j.cub.2017.10.044. [9] MorelliG, EvenA, Gladwyn-NgI, et al. p27Kip1 modulates axonal transport by regulating α-tubulin acetyltransferase 1 stability[J]. Cell Rep, 2018, 23(8): 2429-2442. DOI: 10.1016/j.celrep.2018.04.083. [10] ChawanV, YevateS, GajbhiyeR, et al. Acetylation/deacetylation and microtubule associated proteins influence flagellar axonemal stability and sperm motility[J]. Biosci Rep, 2020, 40(12): BSR20202442. DOI: 10.1042/BSR20202442. [11] DeakinNO, TurnerCE. Paxillin inhibits HDAC6 to regulate microtubule acetylation, Golgi structure, and polarized migration[J]. J Cell Biol, 2014, 206(3): 395-413. DOI: 10.1083/jcb.201403039. [12] NuccitelliR. A role for endogenous electric fields in wound healing[J]. Curr Top Dev Biol, 2003, 58: 1-26. DOI: 10.1016/S0070-2153(03)58001-2. [13] CaiJ, ZhongY, TianS. Naturally occurring davanone terpenoid exhibits anticancer potential against ovarian cancer cells by inducing programmed cell death, by inducing caspase-dependent apoptosis, loss of mitochondrial membrane potential, inhibition of cell migration and invasion and targeting PI3K/AKT/MAPK signaling pathway[J]. J BUON, 2020, 25(5): 2301-2307. [14] GoodsonHV, JonassonEM. Microtubules and microtubule-associated proteins[J]. Cold Spring Harb Perspect Biol, 2018, 10(6): a022608. DOI: 10.1101/cshperspect.a022608. [15] LaFlammeSE, Mathew-SteinerS, SinghN, et al. Integrin and microtubule crosstalk in the regulation of cellular processes[J]. Cell Mol Life Sci, 2018, 75(22): 4177-4185. DOI: 10.1007/s00018-018-2913-x. [16] JankeC, MagieraMM. The tubulin code and its role in controlling microtubule properties and functions[J]. Nat Rev Mol Cell Biol, 2020, 21(6): 307-326. DOI: 10.1038/s41580-020-0214-3. [17] Roll-MecakA. The tubulin code in microtubule dynamics and information encoding[J]. Dev Cell, 2020, 54(1): 7-20. DOI: 10.1016/j.devcel.2020.06.008. [18] LiuN, XiongY, RenY, et al. Proteomic profiling and functional characterization of multiple post-translational modifications of tubulin[J]. J Proteome Res, 2015, 14(8): 3292-3304. DOI: 10.1021/acs.jproteome.5b00308. [19] XuZ, SchaedelL, PortranD, et al. Microtubules acquire resistance from mechanical breakage through intralumenal acetylation[J]. Science, 2017, 356(6335): 328-332. DOI: 10.1126/science.aai8764. [20] Eshun-WilsonL, ZhangR, PortranD, et al. Effects of α-tubulin acetylation on microtubule structure and stability[J]. Proc Natl Acad Sci U S A, 2019, 116(21): 10366-10371. DOI: 10.1073/pnas.1900441116. [21] BanceB, SeetharamanS, LeducC, et al. Microtubule acetylation but not detyrosination promotes focal adhesion dynamics and astrocyte migration[J]. J Cell Sci, 2019, 132(7): jcs225805. DOI: 10.1242/jcs.225805. [22] AtkinsonSJ, GontarczykAM, AlghamdiAA, et al. The β3-integrin endothelial adhesome regulates microtubule-dependent cell migration[J]. EMBO Rep, 2018, 19(7): e44578. DOI: 10.15252/embr.201744578. [23] MyatMM, RashmiRN, MannaD, et al. Drosophila KASH-domain protein Klarsicht regulates microtubule stability and integrin receptor localization during collective cell migration[J]. Dev Biol, 2015, 407(1): 103-114. DOI: 10.1016/j.ydbio.2015.08.003. [24] Rampioni VinciguerraGL, CitronF, SegattoI, et al. p27kip1 at the crossroad between actin and microtubule dynamics[J]. Cell Div, 2019, 14(1): 2. DOI: 10.1186/s13008-019-0045-9. [25] ReedNA, CaiD, BlasiusTL, et al. Microtubule acetylation promotes kinesin-1 binding and transport[J]. Curr Biol, 2006, 16(21): 2166-2172. DOI: 10.1016/j.cub.2006.09.014. [26] Castro-CastroA, JankeC, MontagnacG, et al. ATAT1/MEC-17 acetyltransferase and HDAC6 deacetylase control a balance of acetylation of alpha-tubulin and cortactin and regulate MT1-MMP trafficking and breast tumor cell invasion[J]. Eur J Cell Biol, 2012, 91(11/12): 950-960. DOI: 10.1016/j.ejcb.2012.07.001. [27] van DijkJ, BompardG, CauJ, et al. Microtubule polyglutamylation and acetylation drive microtubule dynamics critical for platelet formation[J]. BMC Biol, 2018, 16(1): 116. DOI: 10.1186/s12915-018-0584-6. [28] ShiP, WangY, HuangY, et al. Arp2/3-branched actin regulates microtubule acetylation levels and affects mitochondrial distribution[J]. J Cell Sci, 2019, 132(6): jcs226506. DOI: 10.1242/jcs.226506. [29] HubbertC, GuardiolaA, ShaoR, et al. HDAC6 is a microtubule-associated deacetylase[J]. Nature, 2002, 417(6887): 455-458. DOI: 10.1038/417455a. [30] AdalbertR, KaiedaA, AntoniouC, et al. Novel HDAC6 inhibitors increase tubulin acetylation and rescue axonal transport of mitochondria in a model of charcot-marie-tooth type 2F[J]. ACS Chem Neurosci, 2020, 11(3): 258-267. DOI: 10.1021/acschemneuro.9b00338. [31] KershawS, MorganDJ, BoydJ, et al. Glucocorticoids rapidly inhibit cell migration through a novel, non-transcriptional HDAC6 pathway[J]. J Cell Sci, 2020, 133(11): jcs242842. DOI: 10.1242/jcs.242842. [32] Valenzuela-FernándezA, CabreroJR, SerradorJM, et al. HDAC6: a key regulator of cytoskeleton, cell migration and cell-cell interactions[J]. Trends Cell Biol, 2008, 18(6): 291-297. DOI: 10.1016/j.tcb.2008.04.003. [33] KeB, ChenY, TuW, et al. Inhibition of HDAC6 activity in kidney diseases: a new perspective[J]. Mol Med, 2018, 24(1): 33. DOI: 10.1186/s10020-018-0027-4. [34] KalinskiAL, KarAN, CraverJ, et al. Deacetylation of Miro1 by HDAC6 blocks mitochondrial transport and mediates axon growth inhibition[J]. J Cell Biol, 2019, 218(6): 1871-1890. DOI: 10.1083/jcb.201702187. [35] SeidelC, SchnekenburgerM, DicatoM, et al. Histone deacetylase 6 in health and disease[J]. Epigenomics, 2015, 7(1): 103-118. DOI: 10.2217/epi.14.69. [36] Shafaq-ZadahM, Gomes-SantosCS, BardinS, et al. Persistent cell migration and adhesion rely on retrograde transport of β1 integrin[J]. Nature Cell Biology, 2016, 18(1): 54-64. DOI: 10.1038/ncb3287. [37] RenX, SunH, LiuJ, et al. Keratinocyte electrotaxis induced by physiological pulsed direct current electric fields[J]. Bioelectrochemistry, 2019, 127: 113-124. DOI: 10.1016/j.bioelechem.2019.02.001. -

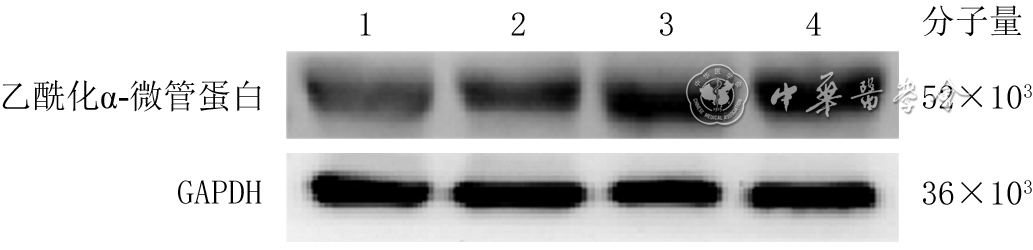
1 蛋白质印迹法检测模拟电场组和用强度200 mV/mm的电场处理不同时间的3组人永生化表皮细胞HaCaT乙酰化α-微管蛋白的表达
注:GAPDH为3-磷酸甘油醛脱氢酶;条带上方1、2、3、4分别为模拟电场组、电场处理1 h组、电场处理2 h 组和电场处理3 h组

2 蛋白质印迹法检测模拟电场组与不同强度电场处理3组人永生化表皮细胞HaCaT中乙酰化α-微管蛋白的表达
注:GAPDH为3-磷酸甘油醛脱氢酶;条带上方1、2、3、4分别为模拟电场组、100 mV/mm电场组、200 mV/mm电场组、300 mV/mm电场组

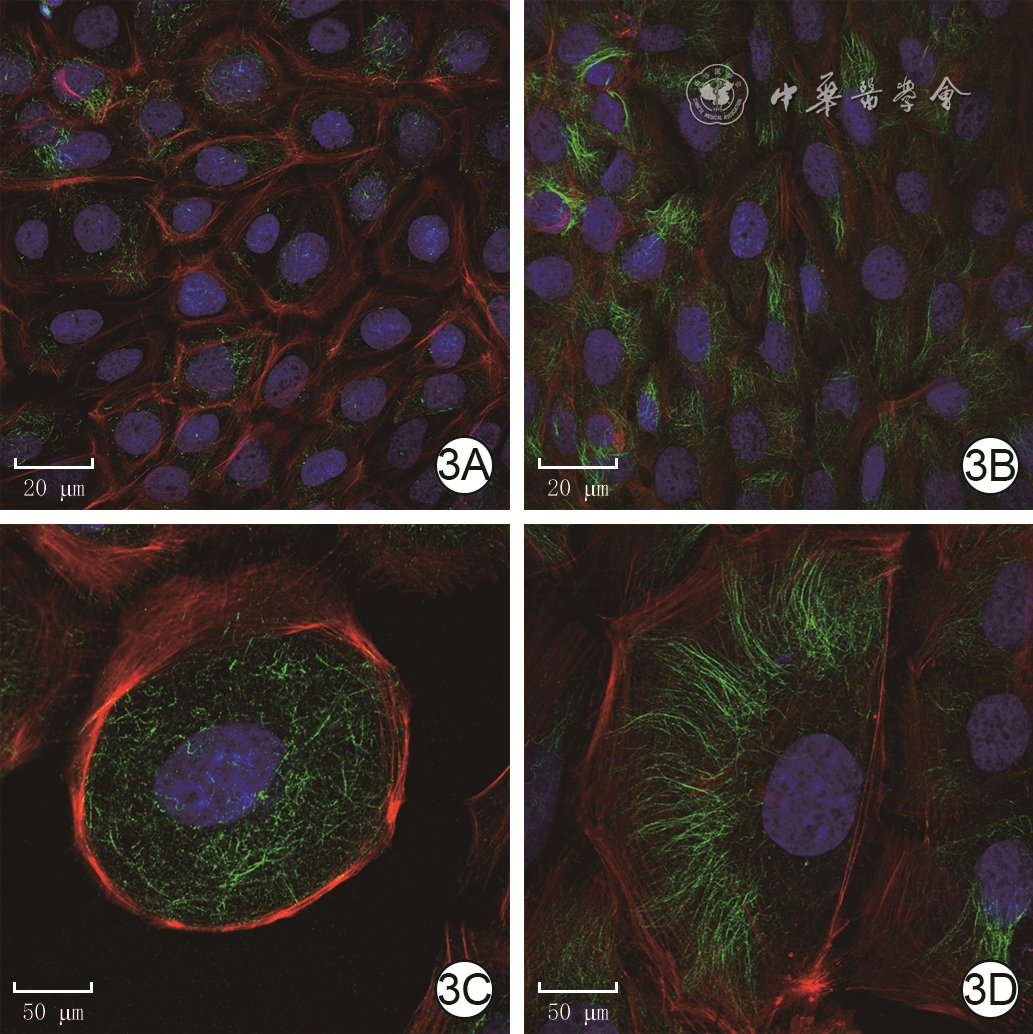
3 模拟电场组和用强度200 mV/mm的电场处理3 h的电场处理组人永生化表皮细胞HaCaT乙酰化α-微管蛋白的表达。3A、3B.分别为模拟电场组和电场处理组HaCaT细胞中乙酰化α-微管蛋白染色重叠图片,图3B中乙酰化α-微管蛋白表达明显多于图3A Alexa Fluor 488-4',6-二脒基-2-苯基吲哚-鬼笔环肽×200;3C、3D.分别为模拟电场组和电场处理组HaCaT细胞中乙酰化α-微管蛋白局部染色重叠图片,图3D中乙酰化α-微管蛋白呈极性分布 Alexa Fluor 488-4',6-二脒基-2-苯基吲哚-鬼笔环肽×600
注:绿色荧光标记乙酰化α-微管蛋白,蓝色荧光标记细胞核,红色荧光标记细胞骨架
表1 2组人永生化表皮细胞HaCaT处理3 h后cosθ与轨迹速度和位移速度比较[M(Q1,Q3)]
组别 样本数 cosθ 轨迹速度(μm/min) 位移速度(μm/min) 模拟电场组 54 0.407(-0.010,0.914) 0.532(0.459,0.641) 0.131(0.068,0.240) 电场处理组 52 -0.986(-0.997,-0.941) 0.628(0.374,0.834) 0.504(0.390,0.548) Z值 -8.65 -2.05 -8.53 P值 <0.001 0.041 <0.001 注:以函数cosθ的计算值量化细胞迁移方向性的变化  下载: 导出CSV
下载: 导出CSV
-

 下载:
下载:



计量
- 文章访问数: 313
- HTML全文浏览量: 105
- PDF下载量: 18
- 被引次数: 0
