



Analysis of genomic information and biological characteristics of a bacteriophage against methicillin-resistant Staphylococcus aureus in patients with median sternal incision infection
-
摘要:
目的 分离提纯1株新型耐甲氧西林金黄色葡萄球菌(MRSA)的噬菌体,并对其基因组学信息和生物学特性进行分析。 方法 采用实验研究方法。取分离自陆军军医大学(第三军医大学)第二附属医院收治的1例胸骨正中切口感染的63岁女性患者创面的MRSA(下称宿主菌)液,采用污水共培养法和双层琼脂平板法从该院污水中分离提纯得到噬菌体,并命名为噬菌体SAP23,观察噬菌斑形态。采用磷钨酸负染法将噬菌体SAP23染色,采用透射电子显微镜观察其形态。采用十二烷基磺酸钠/蛋白酶裂解方案制备噬菌体SAP23 DNA,在Illumina NovaSeq PE150平台下进行全基因组测序,并完成序列组装、注释、系统发生树等基因组学分析。将噬菌体SAP23液分别按10.000 0、1.000 0、0.100 0、0.010 0、0.001 0、0.000 1感染复数与宿主菌液共培养4 h后,采用点滴法测定噬菌体效价,筛选最佳感染复数,此处及以下样本数均为3。按测得的最佳感染复数取噬菌体SAP23液与宿主菌液分别共同孵育5、10、15 min后,同前测定噬菌体效价,筛选最佳吸附时间。按测得的最佳感染复数取噬菌体SAP23液与宿主菌液按最佳吸附时间孵育后,分别于培养0(即刻)、5、10、15、20、30、40、50、60、80、100、120 min,同前测定噬菌体效价,绘制一步生长曲线。取噬菌体SAP23液分别在温度为4、37、50、60、70、80 ℃下,在pH值为2、3、4、5、6、7、8、9、10、11、12下孵育1 h,测定稳定性。取陆军军医大学(第三军医大学)微生物教研室储存的41株MRSA,完成噬菌体SAP23的宿主谱范围检测。 结果 噬菌体SAP23能在宿主菌双层琼脂板上形成透明噬菌斑。噬菌体SAP23头部是直径为(88±4)nm的多面体,其尾部长度为(279±21)nm、宽度为(22.6±2.6)nm。噬菌体SAP23基因组为全长151 618 bp的线状双链DNA,序列两端有11 681 bp的长末端重复序列,预测出220个开放阅读框,噬菌体可编码4个转运RNA,未预测出毒力因子或抗性基因,注释功能的噬菌体SAP23基因可分为5个组,GenBank登录号为MZ427930,噬菌体SAP23全基因组序列与共线性分析中的6个葡萄球菌噬菌体全基因组序列有5个局部共线区域,但在局部共线区域内部或外部存在差异。噬菌体SAP23属于Herelleviridae科Twortvirinae亚科Kayvirus病毒属。噬菌体SAP23的最佳感染复数为0.010 0,最佳吸附时间为10 min,潜伏期约为20 min,裂解期约为80 min;在4~37 ℃温度条件及pH值为4~9的条件中,稳定性较好。噬菌体SAP23可裂解41株MRSA中的3株。 结论 噬菌体SAP23为Herelleviridae科Twortvirinae亚科Kayvirus病毒属成员,潜伏期短,其对温度和酸碱耐受性好,可有效裂解MRSA,为不含毒力因子和抗性基因的新型烈性窄谱噬菌体。 -
关键词:
- 耐甲氧西林金黄色葡萄球菌 /
- 细菌噬菌体 /
- 基因组学 /
- 伤口感染 /
- 生物学特性
Abstract:Objective To isolate and purify a bacteriophage against methicillin-resistant Staphylococcus aureus (MRSA), and to analyze its genomic information and biological characteristics. Methods The experimental research methods were adopted. MRSA (hereinafter referred to as host bacteria) solution was collected from the wound of a 63-year-old female patient with the median sternum incision infection admitted to the Second Affiliated Hospital of Army Medical University (the Third Military Medical University). The bacteriophage, named bacteriophage SAP23 was isolated and purified from the sewage of the Hospital by sewage co-culture method and double-layer agar plate method, and the plaque morphology was observed. The morphology of bacteriophage SAP23 was observed by transmission electron microscope after phosphotungstic acid negative staining. The whole genome of bacteriophage SAP23 was sequenced with NovaSeq PE15 platform after its DNA was prepared by sodium dodecyl sulfonate/protease cleavage scheme, and genomic analysis including sequence assembly, annotation, and phylogenetic tree were completed. The bacteriophage SAP23 solution was co-incubated with the host bacterial solution for 4 h at the multiplicity of infection (MOI) of 10.000 0, 1.000 0, 0.100 0, 0.010 0, 0.001 0, and 0.000 1, respectively, and then the bacteriophage titer was measured by the drip plate method to select the optimal MOI, with here and the following sample numbers of 3. The bacteriophage SAP23 solution was co-incubated with the host bacterial solution at the optimal MOI for 5, 10, and 15 min, respectively, and the bacteriophage titer was measured by the same method as mentioned above to select the optimal adsorption time. After the bacteriophage SAP23 solution was co-incubated with the host bacterial solution at the optimal MOI for the optimal adsorption time, the bacteriophage titers were measured by the same method as mentioned above at 0 (immediately), 5, 10, 15, 20, 30, 40, 50, 60, 80, 100, and 120 min after culture, respectively, and a one-step growth curve was drawn. The bacteriophage SAP23 solution was incubated at 4, 37, 50, 60, 70, and 80 ℃ and pH 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, and 12 for 1 h, respectively, to determine its stability. A total of 41 MRSA strains stored in the Department of Microbiology of Army Medical University (the Third Military Medical University) were used to determine the host spectrum of bacteriophage SAP23. Results The bacteriophage SAP23 could form a transparent plaque on the host bacteria double-layer agar plate. The bacteriophage SAP23 has a polyhedral head with (88±4) nm in diameter and a tail with (279±21) nm in length and (22.6±2.6) nm in width. The bacteriophage SAP23 has a linear, double-stranded DNA with a full length of 151 618 bp and 11 681 bp long terminal repeats sequence in the sequence ends. There were 220 open reading frames predicted and the bacteriophage could encode 4 transfer RNAs, while no resistance genes or virulence factors were found. The annotation function of bacteriophage SAP23 genes could be divided into 5 groups. The GenBank accession number was MZ427930. According to the genomic collinearity analysis, there were 5 local collinear blocks in the whole genome between the bacteriophage SAP23 and the chosen 6 Staphylococcus bacteriophages, while within or outside the local collinear region, there were still some differences. The bacteriophage SAP23 belonged to the Herelleviridae family, Twortvirinae subfamily, and Kayvirus genus. The optimal MOI of bacteriophage SAP23 was 0.010 0, and the optimal adsorption time was 10 min. The bacteriophage SAP23 had a latent period of 20 min, and a growth phase of 80 min. The bacteriophage SAP23 was able to remain stable at the temperature between 4 and 37 ℃ and at the pH values between 4 and 9. The bacteriophage SAP23 could lyse 3 of the 41 tested MRSA strains. Conclusions The bacteriophage SAP23 is a member of the Herelleviridae family, Twortvirinae subfamily, and Kayvirus genus. The bacteriophage SAP23 has a good tolerance for temperature and acid-base and a short latent period, and can lyse MRSA effectively. The bacteriophage SAP23 is a new type of potent narrow-spectrum bacteriophage without virulence factors and resistance genes. -
创面是临床常见病症[1],创面血管新生是创面修复的关键环节之一[2]。血管新生是一个受到精确调控的病理生理过程[3],该过程涉及多种细胞与细胞因子[4, 5],其中微血管内皮细胞是参与此过程的主要功能细胞[6],它们在多种细胞因子及信号通路的精确调控下,通过迁移、增殖、芽生等形成新生血管[7, 8, 9]。
P311,官方基因符号NERP,定位于人5号染色体和小鼠18号染色体上,编码分子量为8×103的胞内蛋白,该蛋白是一种在种属间高度保守的多功能蛋白[10]。研究表明,P311蛋白在组织纤维化[11, 12, 13, 14]、神经系统疾病[15]、肿瘤浸润[16, 17]、血压维持[18]、组织再生[19, 20]等多方面发挥重要作用。近年来研究表明,P311在创面愈合中发挥重要作用[21, 22, 23]。本课题组前期研究表明,人皮肤微血管内皮细胞在静息状态下低表达P311,在炎症刺激条件下高表达P311,进而参与创面血管新生[22,24],但目前P311参与血管新生的机制仍不清楚。因此,本课题组通过前期构建的P311腺病毒转染人微血管内皮细胞1(HMEC-1),使之高表达P311,从而模拟创面愈合过程中P311的表达模式,进而探讨P311对HMEC-1体外血管形成能力的影响及其可能的分子机制。
1. 材料与方法
1.1 主要试剂和仪器来源
基质胶购自美国Corning公司,HMEC系购自美国典型培养物保藏中心,MCDB131培养基购自美国Gibco公司,二甲基亚砜(DMSO)购自德国Meck公司,脂质体2000购自北京索莱宝科技有限公司,细胞计数试剂盒8(CCK-8)、二辛丁酸蛋白浓度测定试剂盒购自海门市碧云天生物技术研究所,VEGF受体2(VEGFR2)、小干扰RNA(siRNA)和阴性对照siRNA均购自广州锐博生物科技有限公司,胞外信号调节激酶1/2(ERK1/2)抑制剂SCH772984购自上海蓝木化工有限公司,兔抗小鼠VEGFR2单克隆抗体、兔抗小鼠磷酸化VEGFR2单克隆抗体、兔抗小鼠ERK1/2单克隆抗体、兔抗小鼠磷酸化ERK1/2单克隆抗体购自美国Cell Signaling Technology公司,小鼠源性GAPDH单克隆抗体购自上海康成生物工程有限公司,辣根过氧化物酶标记的山羊抗兔IgG多克隆抗体和山羊抗小鼠IgG多克隆抗体均购自北京中杉金桥生物技术有限公司。710 型多功能酶标仪购自美国Thermo Fisher公司,LSM510 Meta型活细胞工作站购自德国Zeiss公司,IX71型倒置相差显微镜购自日本Olympus公司,ChemiDoc XRS型凝胶成像仪购自美国Bio-Rad公司。
1.2 HMEC-1的培养和传代
将含有10 ng/mL EGF、10 mmol/L谷氨酰胺、1 µg/mL氢化可的松和体积分数10%胎牛血清的MCDB131培养基(以下称完全培养基)加入底面积25 cm2的培养瓶后,将1×106个HMEC-1接种至该培养瓶中培养,每隔2 d换液1次,待细胞生长达85%融合以上,使用2.5 g/L胰蛋白酶+0.1 g/L乙二胺四乙酸消化后,按1∶3进行细胞传代。取第3~5代细胞用于后续实验。
1.3 P311高表达对HMEC-1增殖、迁移和血管形成能力的影响
1.3.1 细胞分组及处理
用完全培养基调整HMEC-1浓度为1×105个/mL,接种于6孔板中,每孔1 mL。待细胞生长达60%左右融合时,将其按随机数字表法(分组方法下同)分为P311腺病毒组和空载腺病毒组。P311腺病毒组细胞每孔加入10 μL P311腺病毒,空载腺病毒组细胞每孔加入10 μL空载腺病毒(P311腺病毒和空载腺病毒均由本课题组前期构建),转染48 h后,收集细胞用于后续实验。
1.3.2 CCK-8法检测P311高表达对HMEC-1增殖活性的影响
取P311腺病毒组和空载腺病毒组细胞,用完全培养基调整细胞浓度为2×104个/mL,接种于96孔板中,每孔100 μL,每组3个复孔。于培养1、3、5 d,分别使用CCK-8法检测2组细胞增殖活性。检测时,吸弃原有培养基,将完全培养基和CCK-8溶液按体积比10∶1混合,每孔加入混合液100 μL,37 ℃孵育2 h,使用多功能酶标仪测定波长450 nm处的吸光度值,以此代表细胞增殖活性。本实验重复6次。
1.3.3 划痕试验检测P311高表达对HMEC-1迁移能力的影响
取P311腺病毒组和空载腺病毒组细胞,用完全培养基调整细胞浓度为2×104个/mL,接种于24孔板中,每孔1 mL,每组3个复孔。待细胞生长达90%以上融合时,吸弃原有培养基,加入500 μL含2.5 μg/mL丝裂霉素C的完全培养基继续培养2 h。使用100 μL移液枪枪头和直尺在HMEC-1单细胞层上做一直线划痕。吸弃原有培养基,加入PBS冲洗2遍,加入新鲜完全培养基。将细胞置于活细胞工作站倒置相差显微镜200倍镜下观察11 h,记录细胞迁移情况。使用ImageJ 1.48V图像分析软件(美国国立卫生研究院)测量划痕后0(即刻)、6、11 h剩余划痕面积,并计算剩余划痕面积百分比。剩余划痕面积百分比=剩余划痕面积÷初始划痕面积×100%。本实验重复6次。
1.3.4 体外血管形成实验检测P311高表达对HMEC-1血管形成能力的影响
首先将50 μL基质胶加入预冷的96孔板中,在37 ℃培养箱中放置45 min。然后加入用无血清MCDB131培养基培养24 h的P311腺病毒组和空载腺病毒组细胞,用完全培养基调整细胞浓度为4×105个/mL,每孔50 μL,每组3个复孔,在37 ℃培养箱中培养8 h,倒置相差显微镜200倍镜下观察血管形成情况并拍照,采用ImageJ 1.48V图像分析软件测量管状结构节点数和总长度。本实验重复6次。
1.3.5 蛋白质印迹法检测P311高表达对VEGFR2/ERK1/2信号通路的影响
取P311腺病毒组和空载腺病毒组细胞,充分裂解后, 以离心半径10 cm、14 000 r/min离心15 min后收集上清液,二辛丁酸法进行蛋白定量。向上样孔内加入20 μg待测蛋白质样品,十二烷基硫酸钠-聚丙烯酰胺凝胶电泳,湿法转膜后,用30 g/L牛血清白蛋白溶液封闭3 h,分别加入兔抗小鼠VEGFR2单克隆一抗、兔抗小鼠磷酸化VEGFR2单克隆一抗、兔抗小鼠ERK1/2单克隆一抗、兔抗小鼠磷酸化ERK1/2单克隆一抗(稀释比均为1∶1 000)及小鼠源性GAPDH单克隆一抗(稀释比为1∶10 000),4 ℃孵育过夜。次日取出抗体孵育盒,室温下静置1 h,洗涤后分别加入辣根过氧化物酶标记的山羊抗兔IgG多克隆二抗和山羊抗小鼠IgG多克隆二抗(稀释比均为1∶2 000),室温孵育1 h。化学发光液浸泡条带后,凝胶成像仪下曝光,并采集图像,Quantity one软件进行灰度值分析,VEGFR2、ERK1/2的蛋白表达量分别以其与GAPDH灰度值比值表示,磷酸化VEGFR2、磷酸化ERK1/2的蛋白表达量分别以其与VEGFR2、ERK1/2的灰度值比值表示。本实验重复6次。
1.4 P311高表达对HMEC-1 VEGFR2/ERK1/2信号通路及体外血管形成能力的影响
1.4.1 蛋白质印迹法检测siRNA-VEGFR2对P311高表达引起的VEGFR2/ERK1/2信号通路活化的影响
用完全培养基调整HMEC-1浓度为1×105个/mL,接种于6孔板中,每孔1 mL。待细胞生长达60%左右融合时,分为P311腺病毒+阴性对照siRNA组、空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组和空载腺病毒+siRNA-VEGFR2组,每组3个复孔。P311腺病毒+阴性对照siRNA组、空载腺病毒+阴性对照siRNA组细胞每孔分别加入10 μL P311腺病毒、空载腺病毒,24 h后,再采用脂质体2000转染终物质的量浓度50 nmol/L阴性对照siRNA;P311腺病毒+siRNA-VEGFR2组和空载腺病毒+siRNA-VEGFR2组细胞每孔分别加入10 μL P311腺病毒、空载腺病毒,24 h后,采用脂质体2000转染终物质的量浓度50 nmol/L的siRNA-VEGFR2(靶基因序列为5'-GGAAATCTCTTGCAAGCTA-3')。转染24 h,同1.3.5采用蛋白质印迹法检测VEGFR2、磷酸化VEGFR2、ERK1/2、磷酸化ERK1/2的蛋白表达情况。本实验重复6次。
1.4.2 体外血管形成实验检测siRNA-VEGFR2对P311高表达介导的血管形成作用的影响
取HMEC-1,同1.4.1分组及处理,同1.3.4检测各组细胞体外血管形成情况。本实验重复6次。
1.4.3 蛋白质印迹法检测ERK1/2抑制剂对P311高表达引起的ERK1/2信号通路活化的影响
取HMEC-1,用完全培养基调整HMEC-1浓度为1×105个/mL,接种于6孔板中,每孔1 mL,待细胞生长达60%左右融合时,分为P311腺病毒+DMSO组、载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组和空载腺病毒+ERK1/2抑制剂组,每组1孔。P311腺病毒+DMSO组和空载腺病毒+DMSO组细胞每孔分别加入10 μL P311腺病毒、空载腺病毒,24 h后,加入10 μL DMSO处理2 h;P311腺病毒+ERK1/2抑制剂组和空载腺病毒+ERK1/2抑制剂组细胞每孔分别加入10 μL P311腺病毒、空载腺病毒,24 h后,加入终物质的量浓度5 nmol/L ERK1/2抑制剂SCH772984处理2 h。 同1.3.5采用蛋白质印迹法检测ERK1/2和磷酸化ERK1/2的蛋白表达情况。本实验重复6次。
1.4.4 体外血管形成实验检测ERK1/2抑制剂对P311高表达介导的血管形成作用的影响
取HMEC-1,同1.4.3分组及处理,同1.3.4检测各组细胞体外血管形成情况。本实验重复6次。
1.5 统计学处理
采用SPSS 23.0统计软件进行数据分析。所有计量资料数据均符合正态分布,以
2. 结果
2.1 P311高表达对HMEC-1增殖、迁移和血管形成能力的影响
2.1.1 细胞增殖活性
与空载腺病毒组比较,P311腺病毒组细胞培养各时间点增殖活性均没有明显改变(P>0.05),见表1。
表1 2组人微血管内皮细胞1培养各时间点增殖活性比较(组别 样本数 1 d 3 d 5 d P311腺病毒组 6 0.188±0.030 0.415±0.032 0.713±0.030 空载腺病毒组 6 0.185±0.019 0.392±0.030 0.687±0.031 t值 -0.23 -1.30 -1.52 P值 0.822 0.222 0.160 注:处理因素主效应,F=5.10,P=0.074;时间因素主效应,F=720.12,P<0.001;两者交互作用,F=0.81,P=0.473 2.1.2 细胞迁移能力
P311腺病毒组细胞划痕后6、11 h剩余划痕面积百分比较空载腺病毒组明显降低(P<0.05),见表2、图1。
表2 2组人微血管内皮细胞1划痕后各时间点剩余划痕面积百分比比较(%,组别 样本数 6 h 11 h P311腺病毒组 6 47±7 9±5 空载腺病毒组 6 55±5 20±10 t值 -2.47 -2.62 P值 0.033 0.025 注:初始剩余划痕面积百分比均为100%;处理因素主效应,F=18.42,P=0.008;时间因素主效应,F=443.86,P<0.001;两者交互作用,F=7.89,P=0.041  1 2组人微血管内皮细胞1划痕后各时间点剩余划痕面积 倒置相差显微镜×200,图中标尺为100 μm。1A、1B、1C.分别为P311腺病毒组划痕后0(即刻)、6、11 h,随着划痕后时间的延长,剩余划痕面积逐渐减少,至划痕后11 h划痕基本愈合;1D、1E、1F.分别为空载腺病毒组划痕后0、6、11 h,图1E、1F剩余划痕面积分别较图1B、1C明显增大
1 2组人微血管内皮细胞1划痕后各时间点剩余划痕面积 倒置相差显微镜×200,图中标尺为100 μm。1A、1B、1C.分别为P311腺病毒组划痕后0(即刻)、6、11 h,随着划痕后时间的延长,剩余划痕面积逐渐减少,至划痕后11 h划痕基本愈合;1D、1E、1F.分别为空载腺病毒组划痕后0、6、11 h,图1E、1F剩余划痕面积分别较图1B、1C明显增大2.1.3 细胞血管形成能力
培养8 h,P311腺病毒组细胞管状结构节点数为(731±104)个,明显多于空载腺病毒组的(496±74)个(t=4.49,P=0.001);P311腺病毒组细胞管状结构总长度为(19 231±2 647)μm,明显长于空载腺病毒组[(12 670±2 077)μm,t=4.78,P=0.001]。见图2。
 2 2组人微血管内皮细胞1培养8 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。2A. P311腺病毒组管状结构节点数较多,管状结构总长度较长;2B.空载腺病毒组管状结构节点数少于图2A,管状结构总长度短于图2A
2 2组人微血管内皮细胞1培养8 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。2A. P311腺病毒组管状结构节点数较多,管状结构总长度较长;2B.空载腺病毒组管状结构节点数少于图2A,管状结构总长度短于图2A2.1.4 细胞中VEGFR2/ERK1/2信号通路
转染48 h,与空载腺病毒组比较,P311腺病毒组细胞VEGFR2、ERK1/2蛋白表达量均无明显变化(t值分别为0.28、1.36,P值分别为0.786、0.202),磷酸化VEGFR2、磷酸化ERK1/2蛋白表达量均明显升高(t值分别为17.27、16.08,P<0.001)。见图3。
 3 蛋白质印迹法检测2组人微血管内皮细胞1转染48 h VEGFR2/ERK1/2信号通路相关蛋白的表达量。3A.条带图;3B.条图(样本数为6,
3 蛋白质印迹法检测2组人微血管内皮细胞1转染48 h VEGFR2/ERK1/2信号通路相关蛋白的表达量。3A.条带图;3B.条图(样本数为6,2.2 P311通过VEGFR2/ERK1/2信号通路影响HMEC-1体外血管形成能力
2.2.1 siRNA-VEGFR2对P311高表达引起的VEGFR2/ERK1/2信号通路活化的影响
转染24 h,P311腺病毒+阴性对照siRNA组、空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组和空载腺病毒+siRNA-VEGFR2组细胞磷酸化VEGFR2、VEGFR2和磷酸化ERK1/2蛋白表达量总体比较,差异均有统计学意义(F值分别为27.89、25.07、14.20,P<0.001);4组间细胞ERK1/2蛋白表达量总体比较,差异无统计学意义(F=0.08,P=0.971)。
转染24 h,P311腺病毒+阴性对照siRNA组细胞磷酸化VEGFR2和磷酸化ERK1/2蛋白表达量均明显高于空载腺病毒+阴性对照siRNA组(P值分别为<0.001、0.001),2组细胞VEGFR2和ERK1/2蛋白表达量相近(P值分别为0.063、0.640)。P311腺病毒+阴性对照siRNA组细胞VEGFR2、磷酸化VEGFR2、磷酸化ERK1/2蛋白表达量均明显高于P311腺病毒+siRNA-VEGFR2组(P<0.001),2组细胞ERK1/2蛋白表达量相近(P=0.822)。空载腺病毒+阴性对照siRNA组细胞VEGFR2、磷酸化ERK1/2蛋白表达量均明显高于空载腺病毒+siRNA-VEGFR2组(P值分别为<0.001、0.049),该2组细胞磷酸化VEGFR2和ERK1/2蛋白表达量相近(P值分别为0.071、0.761)。转染24 h,P311腺病毒+siRNA-VEGFR2组和空载腺病毒+siRNA-VEGFR2组细胞VEGFR2、磷酸化VEGFR2、ERK1/2和磷酸化ERK1/2蛋白表达量均相近(P值分别为0.155、0.493、0.952、0.314)。见图4。
 4 蛋白质印迹法检测4组人微血管内皮细胞1转染24 h VEGFR2/ERK1/2信号通路相关蛋白的表达量。4A.条带图;4B.条图(样本数为6,
4 蛋白质印迹法检测4组人微血管内皮细胞1转染24 h VEGFR2/ERK1/2信号通路相关蛋白的表达量。4A.条带图;4B.条图(样本数为6,2.2.2 siRNA-VEGFR2对P311高表达介导的血管形成能力的影响
转染24 h,P311腺病毒+阴性对照siRNA组细胞管状结构节点数明显多于空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组(P值均<0.01),管状结构总长度明显长于空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组(P值均<0.01)。转染24 h,空载腺病毒+阴性对照siRNA组细胞管状结构节点数明显多于空载腺病毒+siRNA-VEGFR2组(P<0.01),管状结构总长度明显长于空载腺病毒+siRNA-VEGFR2组(P<0.01)。P311腺病毒+siRNA-VEGFR2组和空载腺病毒+siRNA-VEGFR2组细胞管状结构节点数和管状结构总长度相近(P>0.05)。见图5、表3。
 5 4组人微血管内皮细胞1转染24 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。5A.P311腺病毒+阴性对照小干扰RNA(siRNA)组管状结构节点数较多,管状结构总长度较长;5B.空载腺病毒+阴性对照siRNA组管状结构节点数明显少于图5A,管状结构总长度明显短于图5A;5C.P311腺病毒+siRNA-血管内皮生长因子受体2(VEGFR2)组管状结构节点数明显少于图5A,管状结构总长度明显短于图5A;5D.空载腺病毒+siRNA-VEGFR2组管状结构节点数明显少于图5B,管状结构总长度明显短于图5B,且均与图5C相近表3 4组人微血管内皮细胞1转染24 h后血管形成情况(
5 4组人微血管内皮细胞1转染24 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。5A.P311腺病毒+阴性对照小干扰RNA(siRNA)组管状结构节点数较多,管状结构总长度较长;5B.空载腺病毒+阴性对照siRNA组管状结构节点数明显少于图5A,管状结构总长度明显短于图5A;5C.P311腺病毒+siRNA-血管内皮生长因子受体2(VEGFR2)组管状结构节点数明显少于图5A,管状结构总长度明显短于图5A;5D.空载腺病毒+siRNA-VEGFR2组管状结构节点数明显少于图5B,管状结构总长度明显短于图5B,且均与图5C相近表3 4组人微血管内皮细胞1转染24 h后血管形成情况(组别 样本数 管状结构节点数(个) 管状结构总长度(μm) P311腺病毒+阴性对照siRNA组 6 720±62 21 241±1 139 空载腺病毒+阴性对照siRNA组 6 428±38 17 005±1 156 P311腺病毒+siRNA- VEGFR2组 6 364±57 13 494±2 465 空载腺病毒+siRNA- VEGFR2组 6 310±75 11 600±2 776 F值 57.07 26.32 P值 <0.001 <0.001 P1值 <0.001 0.002 P2值 <0.001 <0.001 P3值 0.002 <0.001 P4值 0.127 0.121 注:F值、P值为4组间各指标总体比较所得;P1值、P2值分别为P311腺病毒+阴性对照小干扰RNA(siRNA)组与空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-血管内皮生长因子受体2(VEGFR2)组比较所得,P3值、P4值分别为空载腺病毒+siRNA-VEGFR2组与空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组比较所得 2.2.3 ERK1/2抑制剂对P311高表达引起的ERK1/2信号通路活化的影响
处理2 h,P311腺病毒+DMSO组、空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组和空载腺病毒+ERK1/2抑制剂组细胞磷酸化ERK1/2蛋白表达量总体比较,差异有统计学意义(F=16.79,P<0.001);4组细胞ERK1/2 蛋白表达量总体比较,差异无统计学意义(F=0.99,P=0.419)。
P311腺病毒+DMSO组细胞磷酸化ERK1/2蛋白表达量明显高于空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组(P值均<0.001),P311腺病毒+DMSO组与空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组细胞ERK1/2蛋白表达量均相近(P值分别为0.806、0.467)。空载腺病毒+DMSO组细胞磷酸化ERK1/2蛋白表达量明显高于空载腺病毒+ERK1/2抑制剂组(P=0.030),2组细胞ERK1/2 蛋白表达量相近(P=0.140)。P311腺病毒+ERK1/2 抑制剂组和空载腺病毒+ERK1/2抑制剂组细胞磷酸化ERK1/2和ERK1/2蛋白表达量均相近(P值分别为0.591、0.373)。见图6。
 6 蛋白质印迹法检测4组人微血管内皮细胞1处理2 h ERK1/2信号通路相关蛋白的表达量。6A.条带图;6B.条图(样本数为6,
6 蛋白质印迹法检测4组人微血管内皮细胞1处理2 h ERK1/2信号通路相关蛋白的表达量。6A.条带图;6B.条图(样本数为6,2.2.4 ERK1/2抑制剂对P311高表达介导的促HMEC-1体外血管形成作用的影响
于处理2 h,P311腺病毒+DMSO组细胞管状结构节点数明显多于空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组(P值均<0.01),管状结构总长度明显长于空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组(P值均<0.01)。空载腺病毒+DMSO组管状结构节点数明显多于空载腺病毒+ERK1/2抑制剂组(P<0.01),管状结构总长度明显长于空载腺病毒+ERK1/2抑制剂组(P<0.01)。P311腺病毒+ERK1/2 抑制剂组和空载腺病毒+ERK1/2抑制剂组细胞管状结构节点数和总长度均相近(P>0.05)。见图7、表4。
 7 4组人微血管内皮细胞1处理2 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。7A.P311腺病毒+二甲基亚砜(DMSO)组管状结构节点数较多,管状结构总长度较长;7B.空载腺病毒+DMSO组管状结构节点数少于图7A,管状结构总长度短于图7A;7C.P311腺病毒+胞外信号调节激酶1/2(ERK1/2)抑制剂组管状结构节点数少于图7A,管状结构总长度短于图7A;7D.空载腺病毒+ERK1/2抑制剂组管状结构节点数和总长度与图7C相近表4 4组人微血管内皮细胞1处理2 h血管形成情况(
7 4组人微血管内皮细胞1处理2 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。7A.P311腺病毒+二甲基亚砜(DMSO)组管状结构节点数较多,管状结构总长度较长;7B.空载腺病毒+DMSO组管状结构节点数少于图7A,管状结构总长度短于图7A;7C.P311腺病毒+胞外信号调节激酶1/2(ERK1/2)抑制剂组管状结构节点数少于图7A,管状结构总长度短于图7A;7D.空载腺病毒+ERK1/2抑制剂组管状结构节点数和总长度与图7C相近表4 4组人微血管内皮细胞1处理2 h血管形成情况(组别 样本数 管状结构节点数(个) 管状结构总长度(μm) P311腺病毒+DMSO组 6 726±72 20 318±1 433 空载腺病毒+DMSO组 6 421±39 16 846±1 464 P311腺病毒+ERK1/2抑制剂组 6 365±41 15 114±1 950 空载腺病毒+ERK1/2抑制剂组 6 317±67 13 188±2 306 F值 63.49 16.53 P值 <0.001 <0.001 P1值 <0.001 0.004 P2值 <0.001 <0.001 P3值 0.005 0.002 P4值 0.162 0.083 注:F值、P值为4组间各指标总体比较所得;DMSO为二甲基亚砜,ERK1/2为胞外信号调节激酶1/2;P1值、P2值分别为P311腺病毒+DMSO组与空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组比较所得,P3值、P4值分别为空载腺病毒+ERK1/2抑制剂组与空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组比较所得 3. 讨论
创面愈合过程被划分为4个连续而又有重叠的阶段,包括凝血期、炎症期、增殖期和重构期[25]。创面一旦形成,就开始了局部新的血液供应系统的建立,该进程贯穿整个创面修复过程[2],但其机制仍不完全清楚。本课题组前期研究表明,皮肤微血管内皮细胞在静息状态下低表达P311,在炎症刺激条件下高表达P311,进而参与创面血管新生[22]。本研究进一步证实,P311能够通过激活VEGFR2/ERK1/2 信号通路,发挥促进HMEC-1血管形成的作用。
P311基因是一种损伤环境诱导型基因,即在静息状态下低表达或者不表达,当受到外界环境刺激时表达升高,从而发挥相关生物学功能[22, 23]。该基因能够编码一种种属间高度保守的细胞内多功能蛋白[10]。在创面愈合过程中,P311持续高表达,可增强表皮干细胞Rho A和Rac1蛋白的活性[21],并激活TGF-β1/Smad信号通路[23],从而发挥促进创面修复作用。近年来研究表明,P311表达于创面皮肤微血管内皮细胞,P311基因敲除导致皮肤创面愈合过程中的血管新生减弱,进而导致创面愈合延迟;P311基因敲除皮肤微血管内皮细胞迁移及血管形成能力均减弱[22]。这些研究结果提示P311可能通过影响微血管内皮细胞的迁移和血管形成能力参与血管新生。微血管内皮细胞具有正常有效的增殖能力在血管形成中也至关重要[26]。
本研究用P311腺病毒转染HMEC-1,CCK-8法检测结果显示,P311腺病毒组和空载腺病毒组HMEC-1增殖能力没有明显差异;划痕试验结果显示,P311腺病毒组HMEC-1迁移能力较空载腺病毒组明显增强;体外血管形成实验结果显示,P311腺病毒组HMEC-1血管形成能力较空载腺病毒组明显增强。以上结果表明P311高表达能够显著提高HMEC-1的迁移能力和体外血管形成能力,而对HMEC-1增殖能力没有明显影响。
VEGFR2信号通路在启动血管内皮细胞各项生物学过程中发挥重要作用[27]。VEGFR2活化是VEGFR2同二聚化或异二聚化导致酪氨酸激酶的活化和受体细胞内结构域中酪氨酸残基的自磷酸化[28, 29]。VEGFR2活化后启动血管内皮细胞内多种信号通路[30]。研究表明,在小鼠中VEGFR2 Tyr1173磷酸化(人类中Tyr1175磷酸化)对ERK1/2 激活至关重要[31]。ERK1/2信号通路与细胞的成活[32]、迁移[33]、血管形成[34]等生物学过程密切相关[35]。然而,此前尚鲜见有关P311对VEGFR2活化和ERK1/2活化的影响的研究。本研究采用蛋白质印迹法检测P311腺病毒组和空载腺病毒组HMEC-1中VEGFR2、磷酸化VEGFR2、ERK1/2和磷酸化ERK1/2的蛋白表达水平,结果显示,P311高表达对HMEC-1 VEGFR2、ERK1/2的蛋白表达没有明显影响,但是却能够促进VEGFR2、ERK1/2的磷酸化。根据上述结果可以得出,P311高表达可以激活VEGFR2/ERK1/2信号通路,该机制可能参与了P311促HMEC-1体外血管形成的作用。
为进一步探讨P311介导的促体外血管形成作用是否通过激活VEGFR2/ERK1/2信号通路而发挥作用,本研究分别采用siRNA-VEGFR2和ERK抑制剂SCH772984阻断VEGFR2/ERK1/2信号通路的活化,随后采用蛋白质印迹法检测细胞VEGFR2磷酸化和ERK1/2磷酸化水平的变化,采用体外血管形成实验检测细胞血管形成能力的变化。结果显示,P311腺病毒+阴性对照siRNA组细胞磷酸化VEGFR2和磷酸化ERK1/2蛋白表达水平均明显高于空载腺病毒+阴性对照siRNA组,提示P311高表达能够促进VEGFR2/ERK1/2信号通路活化。在阻断VEGFR2途径后,VEGFR2和ERK1/2磷酸化水平均明显降低,HMEC-1体外血管形成能力明显减弱,提示siRNA-VEGFR2能够抑制P311高表达引起的VEGFR2/ERK1/2信号通路活化,进而抑制P311高表达介导的促HMEC-1体外血管形成作用。另外,在抑制ERK1/2信号通路后,ERK1/2磷酸化水平均明显降低,HMEC-1体外血管形成能力明显减弱,提示ERK1/2抑制剂能够抑制P311高表达引起的ERK1/2信号通路活化,进而抑制P311高表达介导的促HMEC-1体外血管形成作用。由此推测,P311通过激活VEGFR2/ERK1/2信号通路发挥促进HMEC-1血管形成的作用。
综上所述,P311在创面血管新生中发挥重要作用,能够通过激活VEGFR2/ERK1/2信号通路发挥促进HMEC-1血管形成的作用。本研究结果丰富了创面愈合理论,为促进皮肤创面修复提供了新思路、新线索与新靶点。
·《Burns & Trauma》好文推荐·
严重烫伤后肠道菌群移位的分子机制:囊性纤维化穿膜传导调节蛋白的作用
张建:酝酿和设计实验、实施研究、采集数据、分析/解释数据、文章起草;燕荣帅:酝酿和设计实验、对文章的知识性内容作批评性审阅、指导;杨子晨:酝酿和设计实验、对文章的知识性内容作批评性审阅、获取研究经费、指导、支持性贡献;石茜:实施研究、采集数据;李翔、毛彤春:酝酿和设计实验、采集数据;张一鸣:酝酿和设计实验、文章起草、对文章的知识性内容作批评性审阅、行政、技术或材料支持、指导、支持性贡献所有作者均声明不存在利益冲突 -
参考文献
(63) [1] WangPH, HuangBS, HorngHC, et al.Wound healing[J].J Chin Med Assoc,2018, 81(2):94-101. DOI: 10.1016/j.jcma.2017.11.002. [2] WilkinsonHN, HardmanMJ.Wound healing: cellular mechanisms and pathological outcomes[J].Open Biol,2020, 10(9):200223. DOI: 10.1098/rsob.200223. [3] SarhanWA, AzzazyHME, El-SherbinyIM.Honey/chitosan nanofiber wound dressing enriched with allium sativum and cleome droserifolia: enhanced antimicrobial and wound healing activity[J].ACS Appl Mater Interfaces,2016, 8(10):6379-6390. DOI: 10.1021/acsami.6b00739. [4] HanG, CeilleyR.Chronic wound healing: a review of current management and treatments[J].Adv Ther,2017, 34(3):599-610. DOI: 10.1007/s12325-017-0478-y. [5] SiddiquiAR,BernsteinJM.Chronic wound infection: facts and controversies[J].Clin Dermatol,2010,28(5):519-526.DOI: 10.1016/j.clindermatol.2010.03.009. [6] LakhundiS, ZhangKY. Methicillin-resistant Staphylococcus aureus: molecular characterization, evolution, and epidemiology[J].Clin Microbiol Rev,2018,31(4):e0020-18.DOI: 10.1128/CMR.00020-18. [7] SongRh,YuB,FriedrichD,et al.Naphthoquinone-derivative as a synthetic compound to overcome the antibiotic resistance of methicillin-resistant S. aureus[J].Commun Biol,2020,3(1):529.DOI: 10.1038/s42003-020-01261-0. [8] Hernández-AristizábalI, Ocampo-IbáñezID.Antimicrobial peptides with antibacterial activity against vancomycin-resistant Staphylococcus aureus strains: classification, structures, and mechanisms of action[J].Int J Mol Sci,2021, 22(15):7927. DOI: 10.3390/ijms22157927. [9] CrockerTF, BrownL, LamN, et al. Information provision for stroke survivors and their carers[J].Cochrane Database Syst Rev,2021,11(11):CD001919.DOI: 10.1002/14651858.CD001919.pub4. [10] CisekAA, DąbrowskaI, GregorczykKP, et al.Phage therapy in bacterial infections treatment: one hundred years after the discovery of bacteriophages[J].Curr Microbiol,2017, 74(2):277-283. DOI: 10.1007/s00284-016-1166-x. [11] KortrightKE, ChanBK, KoffJL, et al.Phage therapy: a renewed approach to combat antibiotic-resistant bacteria[J].Cell Host Microbe,2019, 25(2):219-232. DOI: 10.1016/j.chom.2019.01.014. [12] SummersWC.The strange history of phage therapy[J].Bacteriophage,2012, 2(2):130-133. DOI: 10.4161/bact.20757. [13] SarkerSA,SultanaS,ReutelerG,et al. Oral phage therapy of acute bacterial diarrhea with two coliphage preparations: a randomized trial in children from Bangladesh[J].EBioMedicine,2016,4:124-137.DOI: 10.1016/j.ebiom.2015.12.023. [14] FurfaroLL, PayneMS, ChangBJ.Bacteriophage therapy: clinical trials and regulatory hurdles[J].Front Cell Infect Microbiol,2018, 8:376. DOI: 10.3389/fcimb.2018.00376. [15] JaultP,LeclercT,JennesS,et al.Efficacy and tolerability of a cocktail of bacteriophages to treat burn wounds infected by Pseudomonas aeruginosa (PhagoBurn): a randomised, controlled, double-blind phase 1/2 trial[J].Lancet Infect Dis,2019,19(1):35-45.DOI: 10.1016/S1473-3099(18)30482-1. [16] LeitnerL, UjmajuridzeA, ChanishviliN, et al.Intravesical bacteriophages for treating urinary tract infections in patients undergoing transurethral resection of the prostate: a randomised, placebo-controlled, double-blind clinical trial[J].Lancet Infect Dis,2021,21(3):427-436.DOI: 10.1016/S1473-3099(20)30330-3. [17] CarvalhoC, CostaAR, SilvaF, et al.Bacteriophages and their derivatives for the treatment and control of food-producing animal infections[J].Crit Rev Microbiol,2017,43(5):583-601.DOI: 10.1080/1040841X.2016.1271309. [18] MichelsonD, GrundmanM, MagnusonK, et al.Randomized, placebo controlled trial of NPT088, a phage-derived, amyloid-targeted treatment for Alzheimer's disease[J].J Prev Alzheimers Dis,2019, 6(4):228-231. DOI: 10.14283/jpad.2019.37. [19] YangZC, LiuXZ, ShiYL, et al.Characterization and genome annotation of a newly detected bacteriophage infecting multidrug-resistant Acinetobacter baumannii[J].Arch Virol,2019,164(6):1527-1533.DOI: 10.1007/s00705-019-04213-0. [20] LuSG, LeS,TanYL, et al.Genomic and proteomic analyses of the terminally redundant genome of the Pseudomonas aeruginosa phage PaP1: establishment of genus PaP1-like phages[J].PLoS One,2013,8(5):e62933.DOI: 10.1371/journal.pone.0062933. [21] GarneauJR, DepardieuF, FortierLC, et al.PhageTerm: a tool for fast and accurate determination of phage termini and packaging mechanism using next-generation sequencing data[J].Sci Rep,2017,7(1):8292.DOI: 10.1038/s41598-017-07910-5. [22] BesemerJ,LomsadzeA,BorodovskyM.GeneMarkS: a self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions[J].Nucleic Acids Res,2001,29(12):2607-2618.DOI: 10.1093/nar/29.12.2607. [23] BrettinT,DavisJJ,DiszT,et al.RASTtk: a modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes[J].Sci Rep,2015,5:8365.DOI: 10.1038/srep08365. [24] AzizRK,BartelsD,BestAA,et al.The RAST Server: rapid annotations using subsystems technology[J].BMC Genomics,2008,9:75.DOI: 10.1186/1471-2164-9-75. [25] ArndtD,GrantJR,MarcuA,et al. PHASTER: a better, faster version of the PHAST phage search tool[J].Nucleic Acids Res,2016,44(W1):W16-21.DOI: 10.1093/nar/gkw387. [26] LoweTM, EddySR. tRNAscan-SE: a program for improved detection of transfer RNA genes in genomic sequence[J].Nucleic Acids Res,1997,25(5):955-964.DOI: 10.1093/nar/25.5.955. [27] LagesenK,HallinP,RødlandEA,et al.RNAmmer: consistent and rapid annotation of ribosomal RNA genes[J].Nucleic Acids Res,2007,35(9):3100-3108.DOI: 10.1093/nar/gkm160. [28] DarlingACE, MauB, BlattnerFR,et al. Mauve: multiple alignment of conserved genomic sequence with rearrangements[J].Genome Res,2004,14(7):1394-1403.DOI: 10.1101/gr.2289704. [29] AnisimovaM.Evolutionary genomics: statistical and computational methods[M]. New York: Springer New York,2019: 121-147. [30] El-ArabiTF,GriffithsMW,SheYM,et al.Genome sequence and analysis of a broad-host range lytic bacteriophage that infects the Bacillus cereus group[J].Virol J,2013,10:48.DOI: 10.1186/1743-422X-10-48. [31] 吴丽飞,王兆飞,王中华,等.高效裂解多重耐药金黄色葡萄球菌的噬菌体分离及裂解酶的制备[J].中国动物传染病学报,2021,29(3):1-9. DOI: 10.19958/j.cnki.cn31-2031/s.2021.03.001. [32] YangS,YangY,CuiSX,et al.Chitosan-polyvinyl alcohol nanoscale liquid film-forming system facilitates MRSA-infected wound healing by enhancing antibacterial and antibiofilm properties[J].Int J Nanomedicine,2018,13:4987-5002.DOI: 10.2147/IJN.S161680. [33] DouJL, JiangYW, XieJQ, et al.New is old, and old is new: recent advances in antibiotic-based, antibiotic-free and ethnomedical treatments against methicillin-resistant Staphylococcus aureus wound infections[J].Int J Mol Sci,2016, 17(5):617. DOI: 10.3390/ijms17050617. [34] MaciejewskaB,OlszakT,Drulis-KawaZ.Applications of bacteriophages versus phage enzymes to combat and cure bacterial infections: an ambitious and also a realistic application?[J].Appl Microbiol Biotechnol,2018,102(6):2563-2581.DOI: 10.1007/s00253-018-8811-1. [35] MorozovaVV, VlassovVV, TikunovaNV.Applications of bacteriophages in the treatment of localized infections in humans[J].Front Microbiol,2018, 9:1696. DOI: 10.3389/fmicb.2018.01696. [36] OoiML, DrillingAJ, MoralesS, et al.Safety and tolerability of bacteriophage therapy for chronic rhinosinusitis due to Staphylococcus aureus[J].JAMA Otolaryngol Head Neck Surg,2019, 145(8):723-729. DOI: 10.1001/jamaoto.2019.1191. [37] FishR, KutterE, BryanD, et al.Resolving digital staphylococcal osteomyelitis using bacteriophage-a case report[J].Antibiotics (Basel),2018, 7(4):87. DOI: 10.3390/antibiotics7040087. [38] 胡福泉.噬菌体的过去、现在与未来[J].西南医科大学学报,2021,44(5):417-424.DOI: 10.3969/j.issn.2096-3351.2021.05.001. [39] RegeimbalJM, JacobsAC, CoreyBW, et al.Personalized therapeutic cocktail of wild environmental phages rescues mice from Acinetobacter baumannii wound infections[J].Antimicrob Agents Chemother,2016, 60(10):5806-5816. DOI: 10.1128/aac.02877-15. [40] GillJJ, HymanP.Phage choice, isolation, and preparation for phage therapy[J].Curr Pharm Biotechnol,2010, 11(1):2-14. DOI: 10.2174/138920110790725311. [41] KrupovicM,DutilhBE,AdriaenssensEM,et al.Taxonomy of prokaryotic viruses: update from the ICTV bacterial and archaeal viruses subcommittee[J].Arch Virol,2016,161(4):1095-1099.DOI: 10.1007/s00705-015-2728-0. [42] ŁobockaM,HejnowiczMS,DąbrowskiK,et al.Genomics of staphylococcal Twort-like phages--potential therapeutics of the post-antibiotic era[J].Adv Virus Res,2012,83:143-216.DOI: 10.1016/B978-0-12-394438-2.00005-0. [43] AzamAH, TanjiY.Peculiarities of Staphylococcus aureus phages and their possible application in phage therapy[J].Appl Microbiol Biotechnol,2019, 103(11):4279-4289. DOI: 10.1007/s00253-019-09810-2. [44] Głowacka-RutkowskaA, UlatowskaM, EmpelJ, et al.A Kayvirus distant homolog of staphylococcal virulence determinants and VISA biomarker is a phage lytic enzyme[J].Viruses,2020, 12(3):292. DOI: 10.3390/v12030292. [45] DonovanDM,LardeoM,Foster-FreyJ.Lysis of staphylococcal mastitis pathogens by bacteriophage phi11 endolysin[J].FEMS Microbiol Lett,2006,265(1):133-139.DOI: 10.1111/j.1574-6968.2006.00483.x. [46] PaulVD,RajagopalanSS,SundarrajanS,et al.A novel bacteriophage tail-associated muralytic enzyme (TAME) from Phage K and its development into a potent antistaphylococcal protein[J].BMC Microbiol,2011,11:226.DOI: 10.1186/1471-2180-11-226. [47] GuJM,XuW,LeiLC,et al.LysGH15, a novel bacteriophage lysin, protects a murine bacteremia model efficiently against lethal methicillin-resistant Staphylococcus aureus infection[J].J Clin Microbiol,2011,49(1):111-117.DOI: 10.1128/JCM.01144-10. [48] KaurJ, SinghP, SharmaD, et al. A potent enzybiotic against methicillin-resistant Staphylococcus aureus[J].Virus Genes,2020, 56(4):480-497. DOI: 10.1007/s11262-020-01762-4. [49] CahillJ, YoungR.Phage lysis: multiple genes for multiple barriers[J].Adv Virus Res,2019, 103:33-70. DOI: 10.1016/bs.aivir.2018.09.003. [50] LindenSB,ZhangH,HeselpothRD,et al.Biochemical and biophysical characterization of PlyGRCS, a bacteriophage endolysin active against methicillin-resistant Staphylococcus aureus[J].Appl Microbiol Biotechnol,2015,99(2):741-752.DOI: 10.1007/s00253-014-5930-1. [51] 高明明,刘慧莹,李璞媛,等.金黄色葡萄球菌噬菌体vB_SauH_IME522的分离鉴定及全基因组分析[J].第三军医大学学报,2020,42(3):229-240.DOI: 10.16016/j.1000-5404.201909016. [52] ShimamoriY, PramonoAK, KitaoT, et al. Isolation and characterization of a novel phage SaGU1 that infects Staphylococcus aureus clinical isolates from patients with atopic dermatitis[J].Curr Microbiol,2021, 78(4):1267-1276. DOI: 10.1007/s00284-021-02395-y. [53] Bailly-BechetM, VergassolaM, RochaE. Causes for the intriguing presence of tRNAs in phages[J].Genome Res,2007,17(10):1486-1495.DOI: 10.1101/gr.6649807. [54] NunesA, RibeiroDR, MarquesM,et al.Emerging roles of tRNAs in RNA virus infections[J].Trends Biochem Sci,2020,45(9):794-805.DOI: 10.1016/j.tibs.2020.05.007. [55] McCallinS, SarkerSA, BarrettoC,et al.Safety analysis of a Russian phage cocktail: from metagenomic analysis to oral application in healthy human subjects[J].Virology,2013,443(2):187-196.DOI: 10.1016/j.virol.2013.05.022. [56] QuirósP,Colomer-LluchM,Martínez-CastilloA,et al.Antibiotic resistance genes in the bacteriophage DNA fraction of human fecal samples[J].Antimicrob Agents Chemother,2014,58(1):606-609.DOI: 10.1128/AAC.01684-13. [57] 靳晓东,张聪慧,钟江.两株新的金黄色葡萄球菌烈性噬菌体的生物学特性和基因组学研究[J].微生物与感染,2018,13(6):335-341.DOI: 10.3969/j.issn.1673-6184.2018.06.003. [58] JiJW, LiuQ, WangR, et al. Identification of a novel phage targeting methicillin-resistant Staphylococcus aureus in vitro and in vivo[J].Microbial Pathogenesis,2020, 149:104317. DOI: https://doi.org/10.1016/j.micpath.2020.104317. [59] GutiérrezD, VandenheuvelD, MartínezB, et al.Two phages, phiIPLA-RODI and phiIPLA-C1C, lyse mono- and dual-species Staphylococcal biofilms[J].Appl Environ Microbiol,2015, 81(10):3336-3348. DOI: 10.1128/aem.03560-14. [60] FengTT, LeptihnS, DongK, et al. JD419, a Staphylococcus aureus phage with a unique morphology and broad host range[J].Front Microbiol,2021, 12:602902. DOI: 10.3389/fmicb.2021.602902. [61] DoubJB, NgVY, JohnsonAJ, et al.Salvage bacteriophage therapy for a chronic MRSA prosthetic joint infection[J].Antibiotics (Basel),2020, 9(5):241. DOI: 10.3390/antibiotics9050241. [62] JikiaD,ChkhaidzeN,ImedashviliE,et al.The use of a novel biodegradable preparation capable of the sustained release of bacteriophages and ciprofloxacin, in the complex treatment of multidrug-resistant Staphylococcus aureus-infected local radiation injuries caused by exposure to Sr90[J].Clin Exp Dermatol,2005,30(1):23-26.DOI: 10.1111/j.1365-2230.2004.01600.x. [63] LuongT, SalabarriaAC, RoachDR.Phage therapy in the resistance era: where do we stand and where are we going?[J].Clin Ther,2020, 42(9):1659-1680. DOI: 10.1016/j.clinthera.2020.07.014. -

1 从1例胸骨正中切口感染患者创面标本中分离的耐甲氧西林金黄色葡萄球菌噬菌体SAP23及噬菌斑形态。1A.噬菌体SAP23形成透明噬菌斑 图中标尺为10 mm;1B.噬菌体SAP23头部为多面体,并有尾部 透射电子显微镜×40 000,图中标尺为40 nm
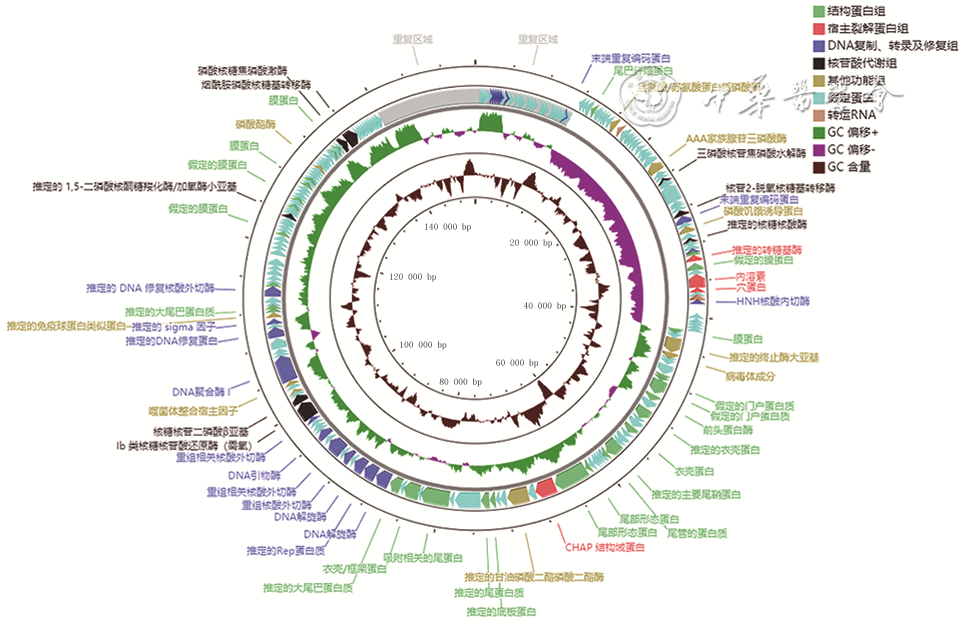
2 从1例胸骨正中切口感染患者创面标本中分离的耐甲氧西林金黄色葡萄球菌噬菌体SAP23基因注释记录图
注:每个箭头代表1个开放阅读框,不同颜色代表编码蛋白不同功能分类;GC为鸟嘌呤胞嘧啶

3 从1例胸骨正中切口感染患者创面标本中分离的耐甲氧西林金黄色葡萄球菌噬菌体SAP23基因组与6株葡萄球菌噬菌体基因组的全基因组共线性记录图
注:1.噬菌体SAP23,2.葡萄球菌噬菌体P108,3.葡萄球菌噬菌体812,4.葡萄球菌噬菌体VB_SavM_JYL02,5.葡萄球菌噬菌体VB_SavM_JYL01,6.葡萄球菌噬菌体vB_SauH_IME522,7.葡萄球菌噬菌体vB_ScoM-PSC1;5种颜色块代表5个局部共线区域;局部共线区域内部或外部的空白区域代表基因组之间的差异区域
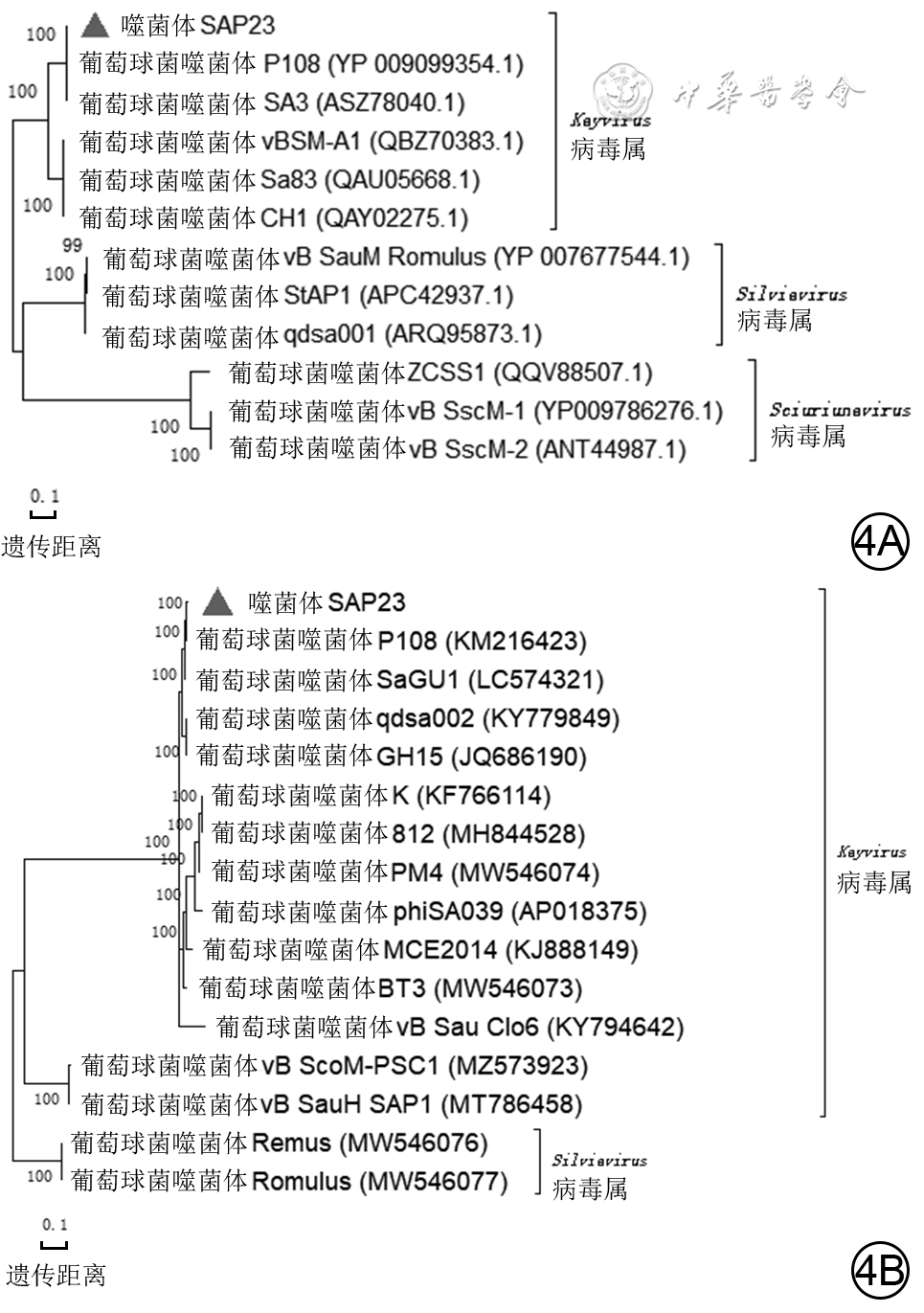
4 从1例胸骨正中切口感染患者创面标本中分离的耐甲氧西林金黄色葡萄球菌噬菌体SAP23系统发生树记录图。4A.主要大衣壳蛋白氨基酸序列系统发生树;4B.全基因组序列系统发生树
注:系统发生树中括号中编号为GenBank登录号,左侧分支数据代表可信度

5 从1例胸骨正中切口感染患者创面标本中分离的耐甲氧西林金黄色葡萄球菌噬菌体SAP23与宿主菌共培养不同时间点的噬菌体效价(
注:PFU为菌斑形成单位;该图为经过lg处理的数据形成的描记图;坐标轴数据为未经lg处理的原始数据
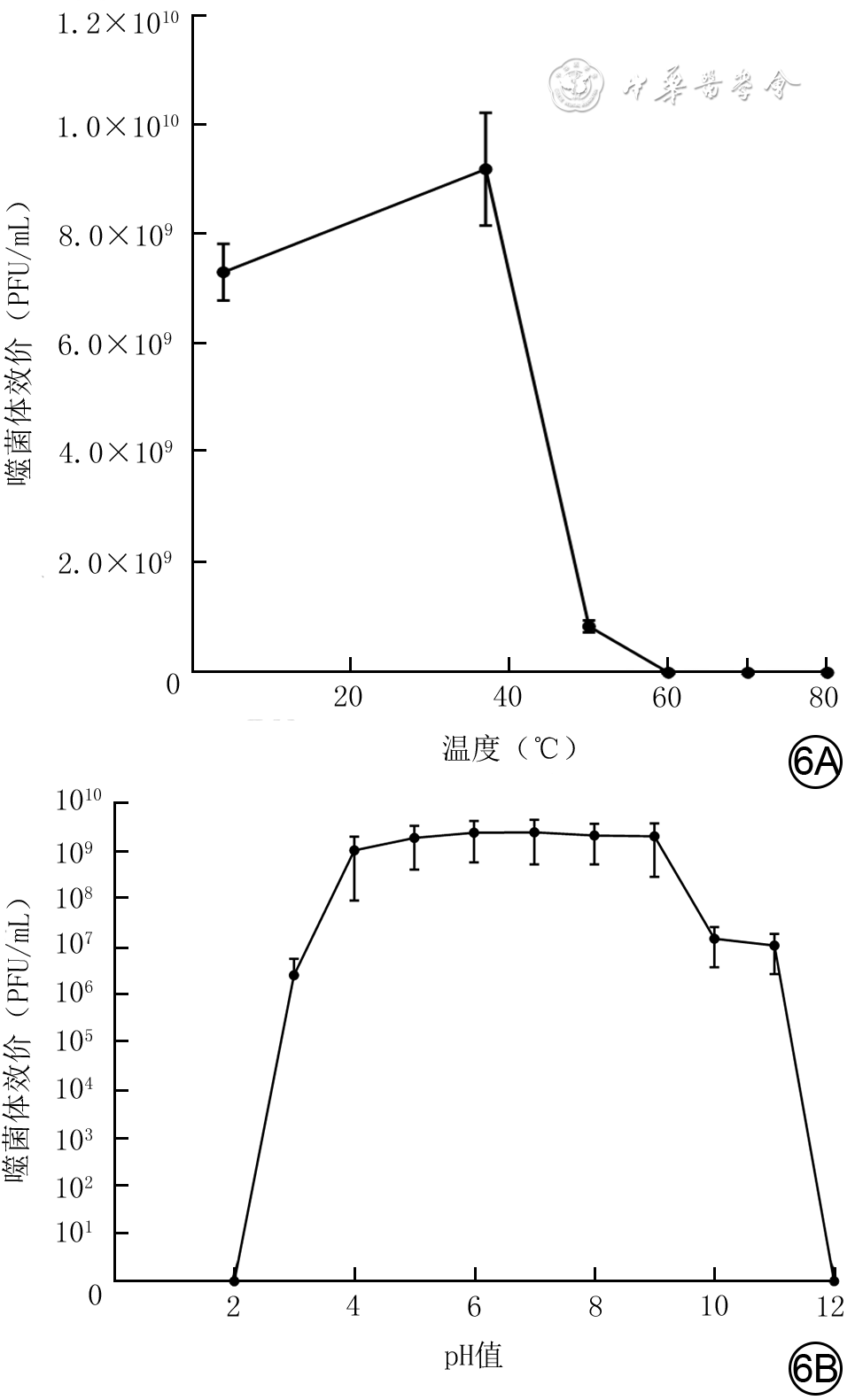
6 从1例胸骨正中切口感染患者创面标本中分离的耐甲氧西林金黄色葡萄球菌噬菌体SAP23在不同温度和pH值条件下的稳定性(
注:PFU为菌斑形成单位;图6B为经过lg处理的数据形成的描记图,坐标轴数据为未经lg处理的原始数据
表1 胸骨正中切口感染患者耐甲氧西林金黄色葡萄球菌噬菌体SAP23对41株耐甲氧西林金黄色葡萄球菌裂解情况
菌株编号 噬菌斑 菌株编号 噬菌斑 菌株编号 噬菌斑 菌株编号 噬菌斑 DP100 - CY6 ++ NF99 - G15 - CY15 - NF7 - SY14 - G26 - CY14 - NF70 - SY19 - G25 - CY19 - NF71 - SY33 - G31 - CY18 - NF73 - SY17 ++ G34 - CY5 - NF75 - SY5 - G13 - CY11 - NF8 - SY13 - G21 - CY16 - NF84 - SY23 - G17 - CY7 - NF88 - SY6 - G36 - CY8 - NF90 + SY15 - G16 - CY20 - 注:“++”表示可见透明噬菌斑,“+”表示可见混浊噬菌斑,“-”表示未见噬菌斑  下载: 导出CSV
下载: 导出CSV
-

 下载:
下载:
计量
- 文章访问数: 884
- HTML全文浏览量: 108
- PDF下载量: 45
- 被引次数: 0
