



Effects of P311 on the angiogenesis ability of human microvascular endothelial cell 1 in vitro and its molecular mechanism
-
摘要:
目的 探讨P311对人微血管内皮细胞1(HMEC-1)血管形成能力的影响及其可能的分子机制。 方法 采用实验研究方法。取HMEC-1,按随机数字表法(分组方法下同)分为P311腺病毒组和空载腺病毒组,分别进行48 h相应转染后,采用细胞计数试剂盒8法检测培养1、3、5 d细胞增殖活性;划痕试验检测细胞划痕后6、11 h剩余划痕面积,并计算剩余划痕面积百分比;体外血管形成实验观察细胞培养8 h血管形成情况,并测量管状结构节点数和总长度;蛋白质印迹法检测细胞中血管内皮生长因子受体2(VEGFR2)、磷酸化VEGFR2、胞外信号调节激酶1/2(ERK1/2)及磷酸化ERK1/2蛋白表达量。取HMEC-1,分为P311腺病毒+阴性对照小干扰RNA(siRNA)组、空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组和空载腺病毒+siRNA-VEGFR2组,分别进行相应的处理,蛋白质印迹法检测转染24 h细胞中VEGFR2、磷酸化VEGFR2、ERK1/2、磷酸化ERK1/2蛋白表达量;体外血管形成实验观察转染24 h细胞血管形成情况,并测量管状结构节点数和总长度。取HMEC-1,分为P311腺病毒+二甲基亚砜(DMSO)组、空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组和空载腺病毒+ERK1/2抑制剂组,分别进行相应的处理。蛋白质印迹法检测处理2 h细胞中ERK1/2及磷酸化ERK1/2蛋白表达量;体外血管形成实验观察处理2 h细胞血管形成情况,并测量管状结构节点数和总长度。各组各时间点样本数均为6。对数据行独立样本t检验、重复测量方差分析、单因素方差分析、LSD检验。 结果 与空载腺病毒组比较,P311腺病毒组细胞培养1、3、5 d增殖活性均没有明显改变(t值分别为-0.23、-1.30、-1.52,P>0.05)。P311腺病毒组细胞划痕后6、11 h剩余划痕面积百分比均较空载腺病毒组明显降低(t值分别为-2.47、-2.62,P<0.05)。培养8 h,与空载腺病毒组比较,P311腺病毒组细胞管状结构节点数和总长度均明显增加(t值分别为4.49、4.78,P<0.01)。转染48 h,与空载腺病毒组比较,P311腺病毒组细胞VEGFR2、ERK1/2蛋白表达量均无明显变化(P>0.05),磷酸化VEGFR2、磷酸化ERK1/2蛋白表达量均明显升高(t值分别为17.27、16.08,P<0.01)。转染24 h,P311腺病毒+阴性对照siRNA组细胞磷酸化VEGFR2和磷酸化ERK1/2蛋白表达量均明显高于空载腺病毒+阴性对照siRNA组(P<0.01),P311腺病毒+阴性对照siRNA组细胞VEGFR2、磷酸化VEGFR2、磷酸化ERK1/2蛋白表达量均明显高于P311腺病毒+siRNA-VEGFR2组(P<0.01),空载腺病毒+阴性对照siRNA组细胞VEGFR2、磷酸化ERK1/2蛋白表达量均明显高于空载腺病毒+siRNA-VEGFR2组(P<0.05或P<0.01)。转染24 h,P311腺病毒+阴性对照siRNA组细胞管状结构节点数为(720±62)个,明显多于空载腺病毒+阴性对照siRNA组的(428±38)个、P311腺病毒+siRNA-VEGFR2组的(364±57)个(P值均<0.01);P311腺病毒+阴性对照siRNA组细胞管状结构总长度为(21 241±1 139)μm,明显长于空载腺病毒+阴性对照siRNA组的(17 005±1 156)μm、P311腺病毒+siRNA-VEGFR2组的(13 494±2 465)μm(P值均<0.01)。空载腺病毒+阴性对照siRNA组细胞管状结构节点数明显多于空载腺病毒+siRNA-VEGFR2组的(310±75)个(P<0.01),管状结构总长度明显长于空载腺病毒+siRNA-VEGFR2组的(11 600±2 776)μm(P<0.01)。处理2 h,P311腺病毒+DMSO组细胞磷酸化ERK1/2蛋白表达量明显高于空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组(P值均<0.01),空载腺病毒+DMSO组细胞磷酸化ERK1/2蛋白表达量明显高于空载腺病毒+ERK1/2抑制剂组(P<0.05)。处理2 h,P311腺病毒+DMSO组细胞管状结构节点数为(726±72)个,明显多于空载腺病毒+DMSO组的(421±39)个、P311腺病毒+ERK1/2抑制剂组的(365±41)个(P值均<0.01);P311腺病毒+DMSO组细胞管状结构总长度为(20 318±1 433)μm,明显长于空载腺病毒+DMSO组的(16 846±1 464)μm、P311腺病毒+ERK1/2抑制剂组的(15 114±1 950)μm(P值均<0.01)。空载腺病毒+DMSO组管状结构节点数明显多于空载腺病毒+ERK1/2抑制剂组的(317±67)个(P<0.01),管状结构总长度明显长于空载腺病毒+ERK1/2抑制剂组的(13 188±2 306)μm(P<0.01)。 结论 P311能够通过激活VEGFR2/ERK1/2信号通路发挥促进HMEC-1血管形成的作用。 -
关键词:
- 伤口愈合 /
- 血管内皮生长因子受体2 /
- 血管新生 /
- 人微血管内皮细胞1 /
- P311 /
- 胞外信号调节激酶1/2
Abstract:Objective To explore the effects of P311 on the angiogenesis ability of human microvascular endothelial cell 1 (HMEC-1) in vitro and the potential molecular mechanism. Methods The experimental research method was used. HMEC-1 was collected and divided into P311 adenovirus group and empty adenovirus group according to the random number table (the same grouping method below), which were transfected correspondingly for 48 h. The cell proliferation activity was detected using the cell counting kit 8 on 1, 3, and 5 days of culture. The residual scratch area of cells at post scratch hour 6 and 11 was detected by scratch test, and the percentage of the residual scratch area was calculated. The blood vessel formation of cells at 8 h of culture was observed by angiogenesis experiment in vitro, and the number of nodes and total length of the tubular structure were measured. The protein expressions of vascular endothelial growth factor receptor 2 (VEGFR2), phosphorylated VEGFR2 (p-VEGFR2), extracellular signal-regulated kinase 1/2 (ERK1/2), and phosphorylated ERK1/2 (p-ERK1/2) in cells were detected by Western blotting. HMEC-1 was collected and divided into P311 adenovirus+small interfering RNA (siRNA) negative control group, empty adenovirus+siRNA negative control group, P311 adenovirus+siRNA-VEGFR2 group, and empty adenovirus+siRNA-VEGFG2 group, which were treated correspondingly. The protein expressions of VEGFR2, p-VEGFR2, ERK1/2, and p-ERK1/2 in cells were detected by Western blotting at 24 h of transfection. The blood vessel formation of cells at 24 h of transfection was observed by angiogenesis experiment in vitro, and the number of nodes and total length of the tubular structure were measured. HMEC-1 was collected and divided into P311 adenovirus+dimethylsulfoxide (DMSO) group, empty adenovirus+DMSO group, P311 adenovirus+ERK1/2 inhibitor group, and empty adenovirus+ERK1/2 inhibitor group, which were treated correspondingly. The protein expressions of ERK1/2 and p-ERK1/2 in cells were detected by Western blotting at 2 h of treatment. The blood vessel formation of cells at 2 h of treatment was observed by angiogenesis experiment in vitro, and the number of nodes and total length of the tubular structure were measured. The sample number at each time point in each group was 6. Data were statistically analyzed with independent sample t test, analysis of variance for repeated measurement, one-way analysis of variance, and least significant difference test. Results Compared with that of empty adenovirus group, the proliferation activity of cells in P311 adenovirus group did not show significant difference on 1, 3, and 5 days of culture (with t values of -0.23, -1.30, and -1.52, respectively, P>0.05). The residual scratch area percentages of cells in P311 adenovirus group were significantly reduced at post scratch hour 6 and 11 compared with those of empty adenovirus group (with t values of -2.47 and -2.62, respectively, P<0.05). At 8 h of culture, compared with those of empty adenovirus group, the number of nodes and total length of the tubular structure of cells in P311 adenovirus group were significantly increased (with t values of 4.49 and 4.78, respectively, P<0.01). At 48 h of transfection, compared with those of empty adenovirus group, the protein expressions of VEGFR2 and ERK1/2 of cells in P311 adenovirus group showed no obvious changes (P>0.05), and the protein expressions of p-VEGFR2 and p-ERK1/2 of cells in P311 adenovirus group were significantly increased (with t values of 17.27 and 16.08, P<0.01). At 24 h of transfection, the protein expressions of p-VEGFR2 and p-ERK1/2 of cells in P311 adenovirus+siRNA negative control group were significantly higher than those in empty adenovirus+siRNA negative control group (P<0.01). The protein expressions of VEGFR2, p-VEGFR2, and p-ERK1/2 of cells in P311 adenovirus+siRNA negative control group were significantly higher than those in P311 adenovirus+siRNA-VEGFR2 group (P<0.01). The protein expressions of VEGFR2 and p-ERK1/2 of cells in empty adenovirus+siRNA negative control group were significantly higher than those in empty adenovirus+siRNA-VEGFR2 group (P<0.05 or P<0.01). At 24 h of transfection, the number of nodes of the tubular structure in cells of P311 adenovirus+siRNA negative control group was 720±62, which was significantly more than 428±38 in empty adenovirus+siRNA negative control group and 364±57 in P311 adenovirus+siRNA-VEGFR2 group (with P values both<0.01). The total length of the tubular structure of cells in P311 adenovirus+siRNA negative control group was (21 241±1 139) μm, which was significantly longer than (17 005±1 156) μm in empty adenovirus+siRNA negative control group and (13 494±2 465) μm in P311 adenovirus+siRNA-VEGFR2 group (with P values both<0.01). The number of nodes of the tubular structure in cells of empty adenovirus+siRNA negative control group was significantly more than 310±75 in empty adenovirus+siRNA-VEGFR2 group (P<0.01), and the total length of the tubular structure of cells in empty adenovirus+siRNA negative control group was significantly longer than (11 600±2 776) μm in empty adenovirus+siRNA-VEGFR2 group (P<0.01). At 2 h of treatment, the protein expression of p-ERK1/2 of cells in P311 adenovirus+DMSO group was significantly higher than that in empty adenovirus+DMSO group and P311 adenovirus+ERK1/2 inhibitor group (with P values both<0.01), and the protein expression of p-ERK1/2 of cells in empty adenovirus+DMSO group was significantly higher than that in empty adenovirus+ERK1/2 inhibitor group (P<0.05). At 2 h of treatment, the number of nodes of the tubular structure in cells of P311 adenovirus+DMSO group was 726±72, which was significantly more than 421±39 in empty adenovirus+DMSO group and 365±41 in P311 adenovirus+ERK1/2 inhibitor group (with P values both<0.01). The total length of the tubular structure of cells in P311 adenovirus+DMSO group was (20 318±1 433) μm, which was significantly longer than (16 846±1 464) μm in empty adenovirus+DMSO group and (15 114±1 950) μm in P311 adenovirus+ERK1/2 inhibitor group (with P values both<0.01). The number of nodes of the tubular structure in cells of empty adenovirus+DMSO group was significantly more than 317±67 in empty adenovirus+ERK1/2 inhibitor group (P<0.01), and the total length of the tubular structure of cells in empty adenovirus+DMSO group was significantly longer than (13 188±2 306) μm in empty adenovirus+ERK1/2 inhibitor group (P<0.01). Conclusions P311 can enhance the angiogenesis ability of HMEC-1 by activating the VEGFR2/ERK1/2 signaling pathway. -
参考文献
(35) [1] FuX.Wound healing center establishment and new technology application in improving the wound healing quality in China[J/OL].Burns Trauma,2020,8:tkaa038[2021-12-10]. https://pubmed.ncbi.nlm.nih.gov/33134399/. DOI: 10.1093/burnst/tkaa038. [2] TonnesenMG,FengX,ClarkRA.Angiogenesis in wound healing[J].J Investig Dermatol Symp Proc,2000,5(1):40-46.DOI: 10.1046/j.1087-0024.2000.00014.x. [3] VeithAP,HendersonK,SpencerA,et al.Therapeutic strategies for enhancing angiogenesis in wound healing[J].Adv Drug Deliv Rev,2019,146:97-125.DOI: 10.1016/j.addr.2018.09.010. [4] FaheyE,DoyleSL.IL-1 family cytokine regulation of vascular permeability and angiogenesis[J].Front Immunol,2019,10:1426.DOI: 10.3389/fimmu.2019.01426. [5] LangmannT.Cytokine signaling as key regulator of pathological angiogenesis in the eye[J].EBioMedicine,2021,73:103662.DOI: 10.1016/j.ebiom.2021.103662. [6] VelnarT,GradisnikL.Tissue augmentation in wound healing: the role of endothelial and epithelial cells[J].Med Arch,2018,72(6):444-448.DOI: 10.5455/medarh.2018.72.444-448. [7] GonzalezAC,CostaTF,AndradeZA,et al.Wound healing- aliterature review[J].An Bras Dermatol,2016,91(5):614-620.DOI: 10.1590/abd1806-4841.20164741. [8] JohnsonKE,WilgusTA.Vascular endothelial growth factor and angiogenesis in the regulation of cutaneous wound repair[J].Adv Wound Care (New Rochelle),2014,3(10):647-661.DOI: 10.1089/wound.2013.0517. [9] GreavesNS,AshcroftKJ,BaguneidM,et al.Current understanding of molecular and cellular mechanisms in fibroplasia and angiogenesis during acute wound healing[J].J Dermatol Sci,2013,72(3):206-217.DOI: 10.1016/j.jdermsci.2013.07.008. [10] WeiZ,HanC,LiH,et al.Molecular mechanism of mesenchyme homeobox 1 in transforming growth factor β1-induced P311 gene transcription in fibrosis[J].Front Mol Biosci,2020,7:59.DOI: 10.3389/fmolb.2020.00059. [11] LagaresD.P311 in scar wars: myofibroblasts lost without transforming growth factor β translation[J].Am J Respir Cell Mol Biol,2019,60(2):139-140.DOI: 10.1165/rcmb.2018-0255ED. [12] YaoZ,YangS,HeW,et al.P311 promotes renal fibrosis via TGFβ1/Smad signaling[J].Sci Rep,2015,5:17032.DOI: 10.1038/srep17032. [13] DuanFF,BarronG,MelitonA,et al.P311 promotes lung fibrosis via stimulation of transforming growth factor-β1, -β2, and -β3 translation[J].Am J Respir Cell Mol Biol,2019,60(2):221-231.DOI: 10.1165/rcmb.2018-0028OC. [14] StradiotL,MannaertsI,van GrunsvenLA.P311, friend, or foe of tissue fibrosis?[J].Front Pharmacol,2018,9:1151.DOI: 10.3389/fphar.2018.01151. [15] TaylorGA,RodriguizRM,GreeneRI,et al.Behavioral characterization of P311 knockout mice[J].Genes Brain Behav,2008,7(7):786-795.DOI: 10.1111/j.1601-183X.2008.00420.x. [16] McDonoughWS,TranNL,BerensME.Regulation of glioma cell migration by serine-phosphorylated P311[J].Neoplasia,2005,7(9):862-872.DOI: 10.1593/neo.05190. [17] KattaK,SembajweLF,Kusche-GullbergM.Potential role for Ext1-dependent heparan sulfate in regulating P311 gene expression in A549 carcinoma cells[J].Biochim Biophys Acta Gen Subj,2018,1862(6):1472-1481.DOI: 10.1016/j.bbagen.2018.03.024. [18] BadriKR,YueM,CarreteroOA,et al.Blood pressure homeostasis is maintained by a P311-TGF-β axis[J].J Clin Invest,2013,123(10):4502-4512.DOI: 10.1172/JCI69884. [19] FujitaniM,YamagishiS,CheYH,et al.P311 accelerates nerve regeneration of the axotomized facial nerve[J].J Neurochem,2004,91(3):737-744.DOI: 10.1111/j.1471-4159.2004.02738.x. [20] ZhaoL,LeungJK,YamamotoH,et al.Identification of P311 as a potential gene regulating alveolar generation[J].Am J Respir Cell Mol Biol,2006,35(1):48-54.DOI: 10.1165/rcmb.2005-0475OC. [21] YaoZ,LiH,HeW,et al.P311 accelerates skin wound reepithelialization by promoting epidermal stem cell migration through RhoA and Rac1 activation[J].Stem Cells Dev,2017,26(6):451-460.DOI: 10.1089/scd.2016.0249. [22] WangS,ZhangX,QianW,et al.P311 deficiency leads to attenuated angiogenesis in cutaneous wound healing[J].Front Physiol,2017,8:1004.DOI: 10.3389/fphys.2017.01004. [23] LiH,YaoZ,HeW,et al.P311 induces the transdifferentiation of epidermal stem cells to myofibroblast-like cells by stimulating transforming growth factor β1 expression[J].Stem Cell Res Ther,2016,7(1):175.DOI: 10.1186/s13287-016-0421-1. [24] ZhouD,LiuT,WangS,et al.Effects of IL-1β and TNF-α on the expression of P311 in vascular endothelial cells and wound healing in mice[J].Front Physiol,2020,11:545008.DOI: 10.3389/fphys.2020.545008. [25] TottoliEM,DoratiR,GentaI,et al.Skin wound healing process and new emerging technologies for skin wound care and regeneration[J].Pharmaceutics,2020,12(8):735. DOI: 10.3390/pharmaceutics12080735. [26] 曹晓赞,谢挺,陆树良.糖化碱性成纤维细胞生长因子影响人真皮微血管内皮细胞增殖和血管化效应的受体途径[J].中华烧伤杂志,2021,37(1):17-24.DOI: 10.3760/cma.j.cn501120-20200916-00412. [27] AbhinandCS,RajuR,SoumyaSJ,et al.VEGF-A/VEGFR2 signaling network in endothelial cells relevant to angiogenesis[J].J Cell Commun Signal,2016,10(4):347-354.DOI: 10.1007/s12079-016-0352-8. [28] LemmonMA,SchlessingerJ.Cell signaling by receptor tyrosine kinases[J].Cell,2010,141(7):1117-1134.DOI: 10.1016/j.cell.2010.06.011. [29] ShibuyaM.VEGFR and type-V RTK activation and signaling[J].Cold Spring Harb Perspect Biol,2013,5(10):a009092.DOI: 10.1101/cshperspect.a009092. [30] SimonsM,GordonE,Claesson-WelshL.Mechanisms and regulation of endothelial VEGF receptor signalling[J].Nat Rev Mol Cell Biol,2016,17(10):611-625.DOI: 10.1038/nrm.2016.87. [31] DellingerMT,BrekkenRA.Phosphorylation of Akt and ERK1/2 is required for VEGF-A/VEGFR2-induced proliferation and migration of lymphatic endothelium[J].PLoS One,2011,6(12):e28947.DOI: 10.1371/journal.pone.0028947. [32] BakerA,WyattD,BocchettaM,et al.Notch-1-PTEN-ERK1/2 signaling axis promotes HER2+ breast cancer cell proliferation and stem cell survival[J].Oncogene,2018,37(33):4489-4504.DOI: 10.1038/s41388-018-0251-y. [33] ValantiEK, Dalakoura-KaragkouniK, FotakisP, et al. Reconstituted HDL-apoE3 promotes endothelial cell migration through ID1 and its downstream kinases ERK1/2, AKT and p38 MAPK[J]. Metabolism, 2022,127:154954. DOI: 10.1016/j.metabol.2021.154954. [34] TangH,HeY,LiL,et al.Exosomal MMP2 derived from mature osteoblasts promotes angiogenesis of endothelial cells via VEGF/Erk1/2 signaling pathway[J].Exp Cell Res,2019,383(2):111541.DOI: 10.1016/j.yexcr.2019.111541. [35] ShinM,BeaneTJ,QuillienA,et al.Vegfa signals through ERK to promote angiogenesis, but not artery differentiation[J].Development,2016,143(20):3796-3805.DOI: 10.1242/dev.137919. -

1 2组人微血管内皮细胞1划痕后各时间点剩余划痕面积 倒置相差显微镜×200,图中标尺为100 μm。1A、1B、1C.分别为P311腺病毒组划痕后0(即刻)、6、11 h,随着划痕后时间的延长,剩余划痕面积逐渐减少,至划痕后11 h划痕基本愈合;1D、1E、1F.分别为空载腺病毒组划痕后0、6、11 h,图1E、1F剩余划痕面积分别较图1B、1C明显增大

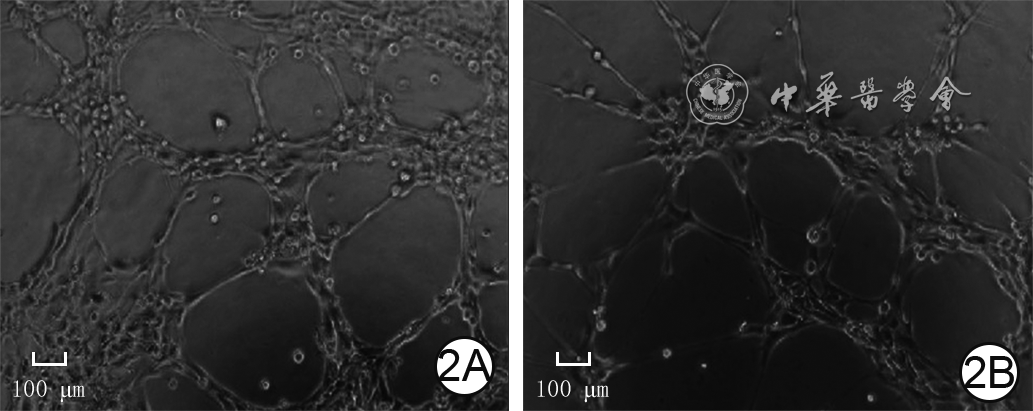
2 2组人微血管内皮细胞1培养8 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。2A. P311腺病毒组管状结构节点数较多,管状结构总长度较长;2B.空载腺病毒组管状结构节点数少于图2A,管状结构总长度短于图2A

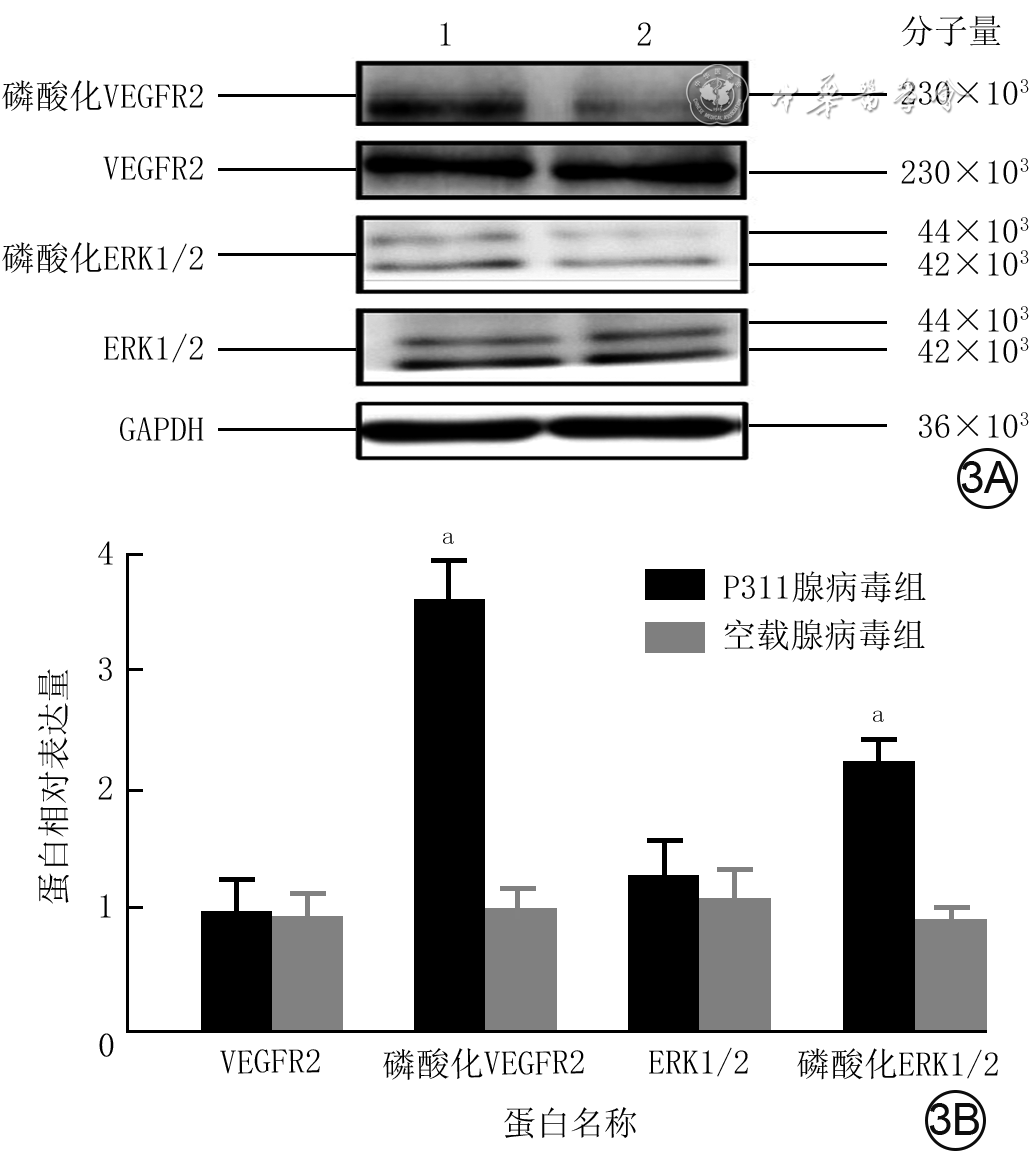
3 蛋白质印迹法检测2组人微血管内皮细胞1转染48 h VEGFR2/ERK1/2信号通路相关蛋白的表达量。3A.条带图;3B.条图(样本数为6,
注:VEGFR2为血管内皮生长因子受体2,ERK1/2为胞外信号调节激酶1/2,GAPDH为3-磷酸甘油醛脱氢酶;条带图上方1、2分别为P311腺病毒组和空载腺病毒组;与空载腺病毒组比较,aP<0.01

4 蛋白质印迹法检测4组人微血管内皮细胞1转染24 h VEGFR2/ERK1/2信号通路相关蛋白的表达量。4A.条带图;4B.条图(样本数为6,
注:VEGFR2为血管内皮生长因子受体2,ERK1/2为胞外信号调节激酶1/2,GAPDH为3-磷酸甘油醛脱氢酶;条带图上方1、2、3、4分别为P311腺病毒+阴性对照小干扰RNA(siRNA)组、空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA- VEGFR2组和空载腺病毒+siRNA-VEGFR2组;与P311腺病毒+阴性对照siRNA组比较,aP<0.01;与空载腺病毒+阴性对照siRNA组比较,bP<0.01,cP<0.05

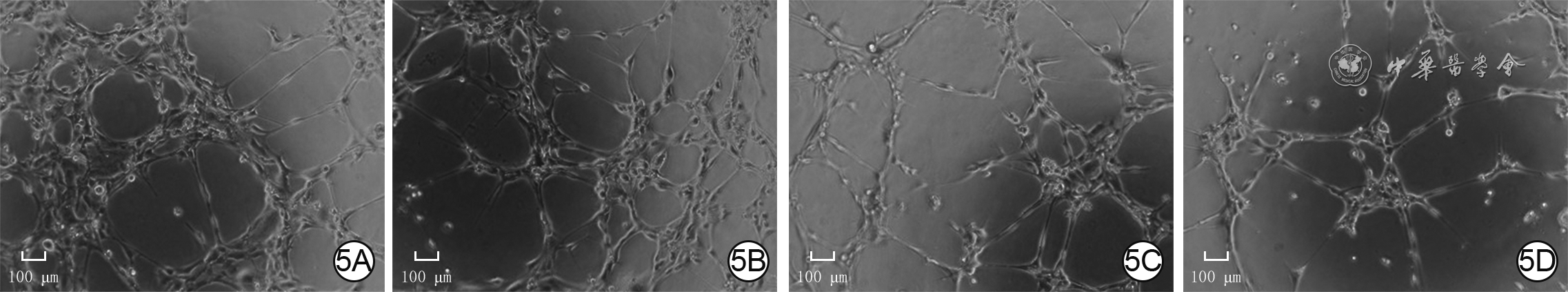
5 4组人微血管内皮细胞1转染24 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。5A.P311腺病毒+阴性对照小干扰RNA(siRNA)组管状结构节点数较多,管状结构总长度较长;5B.空载腺病毒+阴性对照siRNA组管状结构节点数明显少于图5A,管状结构总长度明显短于图5A;5C.P311腺病毒+siRNA-血管内皮生长因子受体2(VEGFR2)组管状结构节点数明显少于图5A,管状结构总长度明显短于图5A;5D.空载腺病毒+siRNA-VEGFR2组管状结构节点数明显少于图5B,管状结构总长度明显短于图5B,且均与图5C相近

6 蛋白质印迹法检测4组人微血管内皮细胞1处理2 h ERK1/2信号通路相关蛋白的表达量。6A.条带图;6B.条图(样本数为6,
注:F值、P值为4组间各指标总体比较所得;ERK1/2为胞外信号调节激酶1/2,GAPDH为3-磷酸甘油醛脱氢酶;条带图上方1、2、3、4分别为P311腺病毒+二甲基亚砜(DMSO)组、空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组和空载腺病毒+ERK1/2抑制剂组;与P311腺病毒+DMSO组比较,aP<0.01;与空载腺病毒+DMSO组比较,bP<0.05

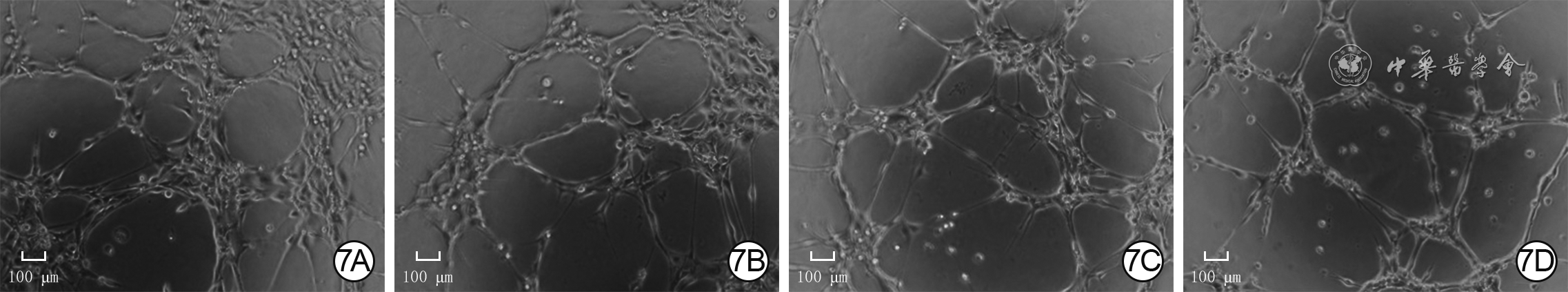
7 4组人微血管内皮细胞1处理2 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。7A.P311腺病毒+二甲基亚砜(DMSO)组管状结构节点数较多,管状结构总长度较长;7B.空载腺病毒+DMSO组管状结构节点数少于图7A,管状结构总长度短于图7A;7C.P311腺病毒+胞外信号调节激酶1/2(ERK1/2)抑制剂组管状结构节点数少于图7A,管状结构总长度短于图7A;7D.空载腺病毒+ERK1/2抑制剂组管状结构节点数和总长度与图7C相近
表1 2组人微血管内皮细胞1培养各时间点增殖活性比较(
组别 样本数 1 d 3 d 5 d P311腺病毒组 6 0.188±0.030 0.415±0.032 0.713±0.030 空载腺病毒组 6 0.185±0.019 0.392±0.030 0.687±0.031 t值 -0.23 -1.30 -1.52 P值 0.822 0.222 0.160 注:处理因素主效应,F=5.10,P=0.074;时间因素主效应,F=720.12,P<0.001;两者交互作用,F=0.81,P=0.473  下载: 导出CSV
下载: 导出CSV
表2 2组人微血管内皮细胞1划痕后各时间点剩余划痕面积百分比比较(%,
组别 样本数 6 h 11 h P311腺病毒组 6 47±7 9±5 空载腺病毒组 6 55±5 20±10 t值 -2.47 -2.62 P值 0.033 0.025 注:初始剩余划痕面积百分比均为100%;处理因素主效应,F=18.42,P=0.008;时间因素主效应,F=443.86,P<0.001;两者交互作用,F=7.89,P=0.041
下载: 导出CSV
表3 4组人微血管内皮细胞1转染24 h后血管形成情况(
组别 样本数 管状结构节点数(个) 管状结构总长度(μm) P311腺病毒+阴性对照siRNA组 6 720±62 21 241±1 139 空载腺病毒+阴性对照siRNA组 6 428±38 17 005±1 156 P311腺病毒+siRNA- VEGFR2组 6 364±57 13 494±2 465 空载腺病毒+siRNA- VEGFR2组 6 310±75 11 600±2 776 F值 57.07 26.32 P值 <0.001 <0.001 P1值 <0.001 0.002 P2值 <0.001 <0.001 P3值 0.002 <0.001 P4值 0.127 0.121 注:F值、P值为4组间各指标总体比较所得;P1值、P2值分别为P311腺病毒+阴性对照小干扰RNA(siRNA)组与空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-血管内皮生长因子受体2(VEGFR2)组比较所得,P3值、P4值分别为空载腺病毒+siRNA-VEGFR2组与空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组比较所得
下载: 导出CSV
表4 4组人微血管内皮细胞1处理2 h血管形成情况(
组别 样本数 管状结构节点数(个) 管状结构总长度(μm) P311腺病毒+DMSO组 6 726±72 20 318±1 433 空载腺病毒+DMSO组 6 421±39 16 846±1 464 P311腺病毒+ERK1/2抑制剂组 6 365±41 15 114±1 950 空载腺病毒+ERK1/2抑制剂组 6 317±67 13 188±2 306 F值 63.49 16.53 P值 <0.001 <0.001 P1值 <0.001 0.004 P2值 <0.001 <0.001 P3值 0.005 0.002 P4值 0.162 0.083 注:F值、P值为4组间各指标总体比较所得;DMSO为二甲基亚砜,ERK1/2为胞外信号调节激酶1/2;P1值、P2值分别为P311腺病毒+DMSO组与空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组比较所得,P3值、P4值分别为空载腺病毒+ERK1/2抑制剂组与空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组比较所得
下载: 导出CSV
-

 下载:
下载:




计量
- 文章访问数: 1118
- HTML全文浏览量: 136
- PDF下载量: 34
- 被引次数: 0
