



Effects of P311 on the angiogenesis ability of human microvascular endothelial cell 1 in vitro and its molecular mechanism
-
摘要:
目的 探讨P311对人微血管内皮细胞1(HMEC-1)血管形成能力的影响及其可能的分子机制。 方法 采用实验研究方法。取HMEC-1,按随机数字表法(分组方法下同)分为P311腺病毒组和空载腺病毒组,分别进行48 h相应转染后,采用细胞计数试剂盒8法检测培养1、3、5 d细胞增殖活性;划痕试验检测细胞划痕后6、11 h剩余划痕面积,并计算剩余划痕面积百分比;体外血管形成实验观察细胞培养8 h血管形成情况,并测量管状结构节点数和总长度;蛋白质印迹法检测细胞中血管内皮生长因子受体2(VEGFR2)、磷酸化VEGFR2、胞外信号调节激酶1/2(ERK1/2)及磷酸化ERK1/2蛋白表达量。取HMEC-1,分为P311腺病毒+阴性对照小干扰RNA(siRNA)组、空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组和空载腺病毒+siRNA-VEGFR2组,分别进行相应的处理,蛋白质印迹法检测转染24 h细胞中VEGFR2、磷酸化VEGFR2、ERK1/2、磷酸化ERK1/2蛋白表达量;体外血管形成实验观察转染24 h细胞血管形成情况,并测量管状结构节点数和总长度。取HMEC-1,分为P311腺病毒+二甲基亚砜(DMSO)组、空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组和空载腺病毒+ERK1/2抑制剂组,分别进行相应的处理。蛋白质印迹法检测处理2 h细胞中ERK1/2及磷酸化ERK1/2蛋白表达量;体外血管形成实验观察处理2 h细胞血管形成情况,并测量管状结构节点数和总长度。各组各时间点样本数均为6。对数据行独立样本t检验、重复测量方差分析、单因素方差分析、LSD检验。 结果 与空载腺病毒组比较,P311腺病毒组细胞培养1、3、5 d增殖活性均没有明显改变(t值分别为-0.23、-1.30、-1.52,P>0.05)。P311腺病毒组细胞划痕后6、11 h剩余划痕面积百分比均较空载腺病毒组明显降低(t值分别为-2.47、-2.62,P<0.05)。培养8 h,与空载腺病毒组比较,P311腺病毒组细胞管状结构节点数和总长度均明显增加(t值分别为4.49、4.78,P<0.01)。转染48 h,与空载腺病毒组比较,P311腺病毒组细胞VEGFR2、ERK1/2蛋白表达量均无明显变化(P>0.05),磷酸化VEGFR2、磷酸化ERK1/2蛋白表达量均明显升高(t值分别为17.27、16.08,P<0.01)。转染24 h,P311腺病毒+阴性对照siRNA组细胞磷酸化VEGFR2和磷酸化ERK1/2蛋白表达量均明显高于空载腺病毒+阴性对照siRNA组(P<0.01),P311腺病毒+阴性对照siRNA组细胞VEGFR2、磷酸化VEGFR2、磷酸化ERK1/2蛋白表达量均明显高于P311腺病毒+siRNA-VEGFR2组(P<0.01),空载腺病毒+阴性对照siRNA组细胞VEGFR2、磷酸化ERK1/2蛋白表达量均明显高于空载腺病毒+siRNA-VEGFR2组(P<0.05或P<0.01)。转染24 h,P311腺病毒+阴性对照siRNA组细胞管状结构节点数为(720±62)个,明显多于空载腺病毒+阴性对照siRNA组的(428±38)个、P311腺病毒+siRNA-VEGFR2组的(364±57)个(P值均<0.01);P311腺病毒+阴性对照siRNA组细胞管状结构总长度为(21 241±1 139)μm,明显长于空载腺病毒+阴性对照siRNA组的(17 005±1 156)μm、P311腺病毒+siRNA-VEGFR2组的(13 494±2 465)μm(P值均<0.01)。空载腺病毒+阴性对照siRNA组细胞管状结构节点数明显多于空载腺病毒+siRNA-VEGFR2组的(310±75)个(P<0.01),管状结构总长度明显长于空载腺病毒+siRNA-VEGFR2组的(11 600±2 776)μm(P<0.01)。处理2 h,P311腺病毒+DMSO组细胞磷酸化ERK1/2蛋白表达量明显高于空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组(P值均<0.01),空载腺病毒+DMSO组细胞磷酸化ERK1/2蛋白表达量明显高于空载腺病毒+ERK1/2抑制剂组(P<0.05)。处理2 h,P311腺病毒+DMSO组细胞管状结构节点数为(726±72)个,明显多于空载腺病毒+DMSO组的(421±39)个、P311腺病毒+ERK1/2抑制剂组的(365±41)个(P值均<0.01);P311腺病毒+DMSO组细胞管状结构总长度为(20 318±1 433)μm,明显长于空载腺病毒+DMSO组的(16 846±1 464)μm、P311腺病毒+ERK1/2抑制剂组的(15 114±1 950)μm(P值均<0.01)。空载腺病毒+DMSO组管状结构节点数明显多于空载腺病毒+ERK1/2抑制剂组的(317±67)个(P<0.01),管状结构总长度明显长于空载腺病毒+ERK1/2抑制剂组的(13 188±2 306)μm(P<0.01)。 结论 P311能够通过激活VEGFR2/ERK1/2信号通路发挥促进HMEC-1血管形成的作用。 -
关键词:
- 伤口愈合 /
- 血管内皮生长因子受体2 /
- 血管新生 /
- 人微血管内皮细胞1 /
- P311 /
- 胞外信号调节激酶1/2
Abstract:Objective To explore the effects of P311 on the angiogenesis ability of human microvascular endothelial cell 1 (HMEC-1) in vitro and the potential molecular mechanism. Methods The experimental research method was used. HMEC-1 was collected and divided into P311 adenovirus group and empty adenovirus group according to the random number table (the same grouping method below), which were transfected correspondingly for 48 h. The cell proliferation activity was detected using the cell counting kit 8 on 1, 3, and 5 days of culture. The residual scratch area of cells at post scratch hour 6 and 11 was detected by scratch test, and the percentage of the residual scratch area was calculated. The blood vessel formation of cells at 8 h of culture was observed by angiogenesis experiment in vitro, and the number of nodes and total length of the tubular structure were measured. The protein expressions of vascular endothelial growth factor receptor 2 (VEGFR2), phosphorylated VEGFR2 (p-VEGFR2), extracellular signal-regulated kinase 1/2 (ERK1/2), and phosphorylated ERK1/2 (p-ERK1/2) in cells were detected by Western blotting. HMEC-1 was collected and divided into P311 adenovirus+small interfering RNA (siRNA) negative control group, empty adenovirus+siRNA negative control group, P311 adenovirus+siRNA-VEGFR2 group, and empty adenovirus+siRNA-VEGFG2 group, which were treated correspondingly. The protein expressions of VEGFR2, p-VEGFR2, ERK1/2, and p-ERK1/2 in cells were detected by Western blotting at 24 h of transfection. The blood vessel formation of cells at 24 h of transfection was observed by angiogenesis experiment in vitro, and the number of nodes and total length of the tubular structure were measured. HMEC-1 was collected and divided into P311 adenovirus+dimethylsulfoxide (DMSO) group, empty adenovirus+DMSO group, P311 adenovirus+ERK1/2 inhibitor group, and empty adenovirus+ERK1/2 inhibitor group, which were treated correspondingly. The protein expressions of ERK1/2 and p-ERK1/2 in cells were detected by Western blotting at 2 h of treatment. The blood vessel formation of cells at 2 h of treatment was observed by angiogenesis experiment in vitro, and the number of nodes and total length of the tubular structure were measured. The sample number at each time point in each group was 6. Data were statistically analyzed with independent sample t test, analysis of variance for repeated measurement, one-way analysis of variance, and least significant difference test. Results Compared with that of empty adenovirus group, the proliferation activity of cells in P311 adenovirus group did not show significant difference on 1, 3, and 5 days of culture (with t values of -0.23, -1.30, and -1.52, respectively, P>0.05). The residual scratch area percentages of cells in P311 adenovirus group were significantly reduced at post scratch hour 6 and 11 compared with those of empty adenovirus group (with t values of -2.47 and -2.62, respectively, P<0.05). At 8 h of culture, compared with those of empty adenovirus group, the number of nodes and total length of the tubular structure of cells in P311 adenovirus group were significantly increased (with t values of 4.49 and 4.78, respectively, P<0.01). At 48 h of transfection, compared with those of empty adenovirus group, the protein expressions of VEGFR2 and ERK1/2 of cells in P311 adenovirus group showed no obvious changes (P>0.05), and the protein expressions of p-VEGFR2 and p-ERK1/2 of cells in P311 adenovirus group were significantly increased (with t values of 17.27 and 16.08, P<0.01). At 24 h of transfection, the protein expressions of p-VEGFR2 and p-ERK1/2 of cells in P311 adenovirus+siRNA negative control group were significantly higher than those in empty adenovirus+siRNA negative control group (P<0.01). The protein expressions of VEGFR2, p-VEGFR2, and p-ERK1/2 of cells in P311 adenovirus+siRNA negative control group were significantly higher than those in P311 adenovirus+siRNA-VEGFR2 group (P<0.01). The protein expressions of VEGFR2 and p-ERK1/2 of cells in empty adenovirus+siRNA negative control group were significantly higher than those in empty adenovirus+siRNA-VEGFR2 group (P<0.05 or P<0.01). At 24 h of transfection, the number of nodes of the tubular structure in cells of P311 adenovirus+siRNA negative control group was 720±62, which was significantly more than 428±38 in empty adenovirus+siRNA negative control group and 364±57 in P311 adenovirus+siRNA-VEGFR2 group (with P values both<0.01). The total length of the tubular structure of cells in P311 adenovirus+siRNA negative control group was (21 241±1 139) μm, which was significantly longer than (17 005±1 156) μm in empty adenovirus+siRNA negative control group and (13 494±2 465) μm in P311 adenovirus+siRNA-VEGFR2 group (with P values both<0.01). The number of nodes of the tubular structure in cells of empty adenovirus+siRNA negative control group was significantly more than 310±75 in empty adenovirus+siRNA-VEGFR2 group (P<0.01), and the total length of the tubular structure of cells in empty adenovirus+siRNA negative control group was significantly longer than (11 600±2 776) μm in empty adenovirus+siRNA-VEGFR2 group (P<0.01). At 2 h of treatment, the protein expression of p-ERK1/2 of cells in P311 adenovirus+DMSO group was significantly higher than that in empty adenovirus+DMSO group and P311 adenovirus+ERK1/2 inhibitor group (with P values both<0.01), and the protein expression of p-ERK1/2 of cells in empty adenovirus+DMSO group was significantly higher than that in empty adenovirus+ERK1/2 inhibitor group (P<0.05). At 2 h of treatment, the number of nodes of the tubular structure in cells of P311 adenovirus+DMSO group was 726±72, which was significantly more than 421±39 in empty adenovirus+DMSO group and 365±41 in P311 adenovirus+ERK1/2 inhibitor group (with P values both<0.01). The total length of the tubular structure of cells in P311 adenovirus+DMSO group was (20 318±1 433) μm, which was significantly longer than (16 846±1 464) μm in empty adenovirus+DMSO group and (15 114±1 950) μm in P311 adenovirus+ERK1/2 inhibitor group (with P values both<0.01). The number of nodes of the tubular structure in cells of empty adenovirus+DMSO group was significantly more than 317±67 in empty adenovirus+ERK1/2 inhibitor group (P<0.01), and the total length of the tubular structure of cells in empty adenovirus+DMSO group was significantly longer than (13 188±2 306) μm in empty adenovirus+ERK1/2 inhibitor group (P<0.01). Conclusions P311 can enhance the angiogenesis ability of HMEC-1 by activating the VEGFR2/ERK1/2 signaling pathway. -
创面是临床常见病症[1],创面血管新生是创面修复的关键环节之一[2]。血管新生是一个受到精确调控的病理生理过程[3],该过程涉及多种细胞与细胞因子[4, 5],其中微血管内皮细胞是参与此过程的主要功能细胞[6],它们在多种细胞因子及信号通路的精确调控下,通过迁移、增殖、芽生等形成新生血管[7, 8, 9]。
P311,官方基因符号NERP,定位于人5号染色体和小鼠18号染色体上,编码分子量为8×103的胞内蛋白,该蛋白是一种在种属间高度保守的多功能蛋白[10]。研究表明,P311蛋白在组织纤维化[11, 12, 13, 14]、神经系统疾病[15]、肿瘤浸润[16, 17]、血压维持[18]、组织再生[19, 20]等多方面发挥重要作用。近年来研究表明,P311在创面愈合中发挥重要作用[21, 22, 23]。本课题组前期研究表明,人皮肤微血管内皮细胞在静息状态下低表达P311,在炎症刺激条件下高表达P311,进而参与创面血管新生[22,24],但目前P311参与血管新生的机制仍不清楚。因此,本课题组通过前期构建的P311腺病毒转染人微血管内皮细胞1(HMEC-1),使之高表达P311,从而模拟创面愈合过程中P311的表达模式,进而探讨P311对HMEC-1体外血管形成能力的影响及其可能的分子机制。
1. 材料与方法
1.1 主要试剂和仪器来源
基质胶购自美国Corning公司,HMEC系购自美国典型培养物保藏中心,MCDB131培养基购自美国Gibco公司,二甲基亚砜(DMSO)购自德国Meck公司,脂质体2000购自北京索莱宝科技有限公司,细胞计数试剂盒8(CCK-8)、二辛丁酸蛋白浓度测定试剂盒购自海门市碧云天生物技术研究所,VEGF受体2(VEGFR2)、小干扰RNA(siRNA)和阴性对照siRNA均购自广州锐博生物科技有限公司,胞外信号调节激酶1/2(ERK1/2)抑制剂SCH772984购自上海蓝木化工有限公司,兔抗小鼠VEGFR2单克隆抗体、兔抗小鼠磷酸化VEGFR2单克隆抗体、兔抗小鼠ERK1/2单克隆抗体、兔抗小鼠磷酸化ERK1/2单克隆抗体购自美国Cell Signaling Technology公司,小鼠源性GAPDH单克隆抗体购自上海康成生物工程有限公司,辣根过氧化物酶标记的山羊抗兔IgG多克隆抗体和山羊抗小鼠IgG多克隆抗体均购自北京中杉金桥生物技术有限公司。710 型多功能酶标仪购自美国Thermo Fisher公司,LSM510 Meta型活细胞工作站购自德国Zeiss公司,IX71型倒置相差显微镜购自日本Olympus公司,ChemiDoc XRS型凝胶成像仪购自美国Bio-Rad公司。
1.2 HMEC-1的培养和传代
将含有10 ng/mL EGF、10 mmol/L谷氨酰胺、1 µg/mL氢化可的松和体积分数10%胎牛血清的MCDB131培养基(以下称完全培养基)加入底面积25 cm2的培养瓶后,将1×106个HMEC-1接种至该培养瓶中培养,每隔2 d换液1次,待细胞生长达85%融合以上,使用2.5 g/L胰蛋白酶+0.1 g/L乙二胺四乙酸消化后,按1∶3进行细胞传代。取第3~5代细胞用于后续实验。
1.3 P311高表达对HMEC-1增殖、迁移和血管形成能力的影响
1.3.1 细胞分组及处理
用完全培养基调整HMEC-1浓度为1×105个/mL,接种于6孔板中,每孔1 mL。待细胞生长达60%左右融合时,将其按随机数字表法(分组方法下同)分为P311腺病毒组和空载腺病毒组。P311腺病毒组细胞每孔加入10 μL P311腺病毒,空载腺病毒组细胞每孔加入10 μL空载腺病毒(P311腺病毒和空载腺病毒均由本课题组前期构建),转染48 h后,收集细胞用于后续实验。
1.3.2 CCK-8法检测P311高表达对HMEC-1增殖活性的影响
取P311腺病毒组和空载腺病毒组细胞,用完全培养基调整细胞浓度为2×104个/mL,接种于96孔板中,每孔100 μL,每组3个复孔。于培养1、3、5 d,分别使用CCK-8法检测2组细胞增殖活性。检测时,吸弃原有培养基,将完全培养基和CCK-8溶液按体积比10∶1混合,每孔加入混合液100 μL,37 ℃孵育2 h,使用多功能酶标仪测定波长450 nm处的吸光度值,以此代表细胞增殖活性。本实验重复6次。
1.3.3 划痕试验检测P311高表达对HMEC-1迁移能力的影响
取P311腺病毒组和空载腺病毒组细胞,用完全培养基调整细胞浓度为2×104个/mL,接种于24孔板中,每孔1 mL,每组3个复孔。待细胞生长达90%以上融合时,吸弃原有培养基,加入500 μL含2.5 μg/mL丝裂霉素C的完全培养基继续培养2 h。使用100 μL移液枪枪头和直尺在HMEC-1单细胞层上做一直线划痕。吸弃原有培养基,加入PBS冲洗2遍,加入新鲜完全培养基。将细胞置于活细胞工作站倒置相差显微镜200倍镜下观察11 h,记录细胞迁移情况。使用ImageJ 1.48V图像分析软件(美国国立卫生研究院)测量划痕后0(即刻)、6、11 h剩余划痕面积,并计算剩余划痕面积百分比。剩余划痕面积百分比=剩余划痕面积÷初始划痕面积×100%。本实验重复6次。
1.3.4 体外血管形成实验检测P311高表达对HMEC-1血管形成能力的影响
首先将50 μL基质胶加入预冷的96孔板中,在37 ℃培养箱中放置45 min。然后加入用无血清MCDB131培养基培养24 h的P311腺病毒组和空载腺病毒组细胞,用完全培养基调整细胞浓度为4×105个/mL,每孔50 μL,每组3个复孔,在37 ℃培养箱中培养8 h,倒置相差显微镜200倍镜下观察血管形成情况并拍照,采用ImageJ 1.48V图像分析软件测量管状结构节点数和总长度。本实验重复6次。
1.3.5 蛋白质印迹法检测P311高表达对VEGFR2/ERK1/2信号通路的影响
取P311腺病毒组和空载腺病毒组细胞,充分裂解后, 以离心半径10 cm、14 000 r/min离心15 min后收集上清液,二辛丁酸法进行蛋白定量。向上样孔内加入20 μg待测蛋白质样品,十二烷基硫酸钠-聚丙烯酰胺凝胶电泳,湿法转膜后,用30 g/L牛血清白蛋白溶液封闭3 h,分别加入兔抗小鼠VEGFR2单克隆一抗、兔抗小鼠磷酸化VEGFR2单克隆一抗、兔抗小鼠ERK1/2单克隆一抗、兔抗小鼠磷酸化ERK1/2单克隆一抗(稀释比均为1∶1 000)及小鼠源性GAPDH单克隆一抗(稀释比为1∶10 000),4 ℃孵育过夜。次日取出抗体孵育盒,室温下静置1 h,洗涤后分别加入辣根过氧化物酶标记的山羊抗兔IgG多克隆二抗和山羊抗小鼠IgG多克隆二抗(稀释比均为1∶2 000),室温孵育1 h。化学发光液浸泡条带后,凝胶成像仪下曝光,并采集图像,Quantity one软件进行灰度值分析,VEGFR2、ERK1/2的蛋白表达量分别以其与GAPDH灰度值比值表示,磷酸化VEGFR2、磷酸化ERK1/2的蛋白表达量分别以其与VEGFR2、ERK1/2的灰度值比值表示。本实验重复6次。
1.4 P311高表达对HMEC-1 VEGFR2/ERK1/2信号通路及体外血管形成能力的影响
1.4.1 蛋白质印迹法检测siRNA-VEGFR2对P311高表达引起的VEGFR2/ERK1/2信号通路活化的影响
用完全培养基调整HMEC-1浓度为1×105个/mL,接种于6孔板中,每孔1 mL。待细胞生长达60%左右融合时,分为P311腺病毒+阴性对照siRNA组、空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组和空载腺病毒+siRNA-VEGFR2组,每组3个复孔。P311腺病毒+阴性对照siRNA组、空载腺病毒+阴性对照siRNA组细胞每孔分别加入10 μL P311腺病毒、空载腺病毒,24 h后,再采用脂质体2000转染终物质的量浓度50 nmol/L阴性对照siRNA;P311腺病毒+siRNA-VEGFR2组和空载腺病毒+siRNA-VEGFR2组细胞每孔分别加入10 μL P311腺病毒、空载腺病毒,24 h后,采用脂质体2000转染终物质的量浓度50 nmol/L的siRNA-VEGFR2(靶基因序列为5'-GGAAATCTCTTGCAAGCTA-3')。转染24 h,同1.3.5采用蛋白质印迹法检测VEGFR2、磷酸化VEGFR2、ERK1/2、磷酸化ERK1/2的蛋白表达情况。本实验重复6次。
1.4.2 体外血管形成实验检测siRNA-VEGFR2对P311高表达介导的血管形成作用的影响
取HMEC-1,同1.4.1分组及处理,同1.3.4检测各组细胞体外血管形成情况。本实验重复6次。
1.4.3 蛋白质印迹法检测ERK1/2抑制剂对P311高表达引起的ERK1/2信号通路活化的影响
取HMEC-1,用完全培养基调整HMEC-1浓度为1×105个/mL,接种于6孔板中,每孔1 mL,待细胞生长达60%左右融合时,分为P311腺病毒+DMSO组、载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组和空载腺病毒+ERK1/2抑制剂组,每组1孔。P311腺病毒+DMSO组和空载腺病毒+DMSO组细胞每孔分别加入10 μL P311腺病毒、空载腺病毒,24 h后,加入10 μL DMSO处理2 h;P311腺病毒+ERK1/2抑制剂组和空载腺病毒+ERK1/2抑制剂组细胞每孔分别加入10 μL P311腺病毒、空载腺病毒,24 h后,加入终物质的量浓度5 nmol/L ERK1/2抑制剂SCH772984处理2 h。 同1.3.5采用蛋白质印迹法检测ERK1/2和磷酸化ERK1/2的蛋白表达情况。本实验重复6次。
1.4.4 体外血管形成实验检测ERK1/2抑制剂对P311高表达介导的血管形成作用的影响
取HMEC-1,同1.4.3分组及处理,同1.3.4检测各组细胞体外血管形成情况。本实验重复6次。
1.5 统计学处理
采用SPSS 23.0统计软件进行数据分析。所有计量资料数据均符合正态分布,以
2. 结果
2.1 P311高表达对HMEC-1增殖、迁移和血管形成能力的影响
2.1.1 细胞增殖活性
与空载腺病毒组比较,P311腺病毒组细胞培养各时间点增殖活性均没有明显改变(P>0.05),见表1。
表1 2组人微血管内皮细胞1培养各时间点增殖活性比较(组别 样本数 1 d 3 d 5 d P311腺病毒组 6 0.188±0.030 0.415±0.032 0.713±0.030 空载腺病毒组 6 0.185±0.019 0.392±0.030 0.687±0.031 t值 -0.23 -1.30 -1.52 P值 0.822 0.222 0.160 注:处理因素主效应,F=5.10,P=0.074;时间因素主效应,F=720.12,P<0.001;两者交互作用,F=0.81,P=0.473 2.1.2 细胞迁移能力
P311腺病毒组细胞划痕后6、11 h剩余划痕面积百分比较空载腺病毒组明显降低(P<0.05),见表2、图1。
表2 2组人微血管内皮细胞1划痕后各时间点剩余划痕面积百分比比较(%,组别 样本数 6 h 11 h P311腺病毒组 6 47±7 9±5 空载腺病毒组 6 55±5 20±10 t值 -2.47 -2.62 P值 0.033 0.025 注:初始剩余划痕面积百分比均为100%;处理因素主效应,F=18.42,P=0.008;时间因素主效应,F=443.86,P<0.001;两者交互作用,F=7.89,P=0.041  1 2组人微血管内皮细胞1划痕后各时间点剩余划痕面积 倒置相差显微镜×200,图中标尺为100 μm。1A、1B、1C.分别为P311腺病毒组划痕后0(即刻)、6、11 h,随着划痕后时间的延长,剩余划痕面积逐渐减少,至划痕后11 h划痕基本愈合;1D、1E、1F.分别为空载腺病毒组划痕后0、6、11 h,图1E、1F剩余划痕面积分别较图1B、1C明显增大
1 2组人微血管内皮细胞1划痕后各时间点剩余划痕面积 倒置相差显微镜×200,图中标尺为100 μm。1A、1B、1C.分别为P311腺病毒组划痕后0(即刻)、6、11 h,随着划痕后时间的延长,剩余划痕面积逐渐减少,至划痕后11 h划痕基本愈合;1D、1E、1F.分别为空载腺病毒组划痕后0、6、11 h,图1E、1F剩余划痕面积分别较图1B、1C明显增大2.1.3 细胞血管形成能力
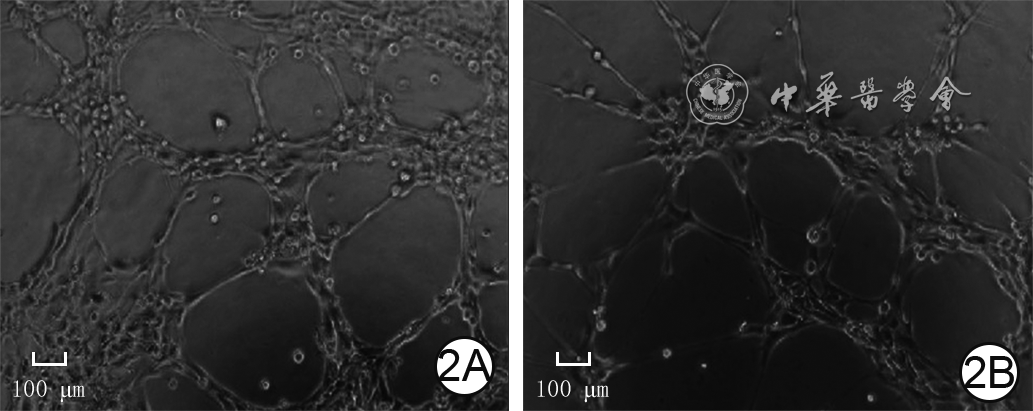
培养8 h,P311腺病毒组细胞管状结构节点数为(731±104)个,明显多于空载腺病毒组的(496±74)个(t=4.49,P=0.001);P311腺病毒组细胞管状结构总长度为(19 231±2 647)μm,明显长于空载腺病毒组[(12 670±2 077)μm,t=4.78,P=0.001]。见图2。
 2 2组人微血管内皮细胞1培养8 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。2A. P311腺病毒组管状结构节点数较多,管状结构总长度较长;2B.空载腺病毒组管状结构节点数少于图2A,管状结构总长度短于图2A
2 2组人微血管内皮细胞1培养8 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。2A. P311腺病毒组管状结构节点数较多,管状结构总长度较长;2B.空载腺病毒组管状结构节点数少于图2A,管状结构总长度短于图2A2.1.4 细胞中VEGFR2/ERK1/2信号通路
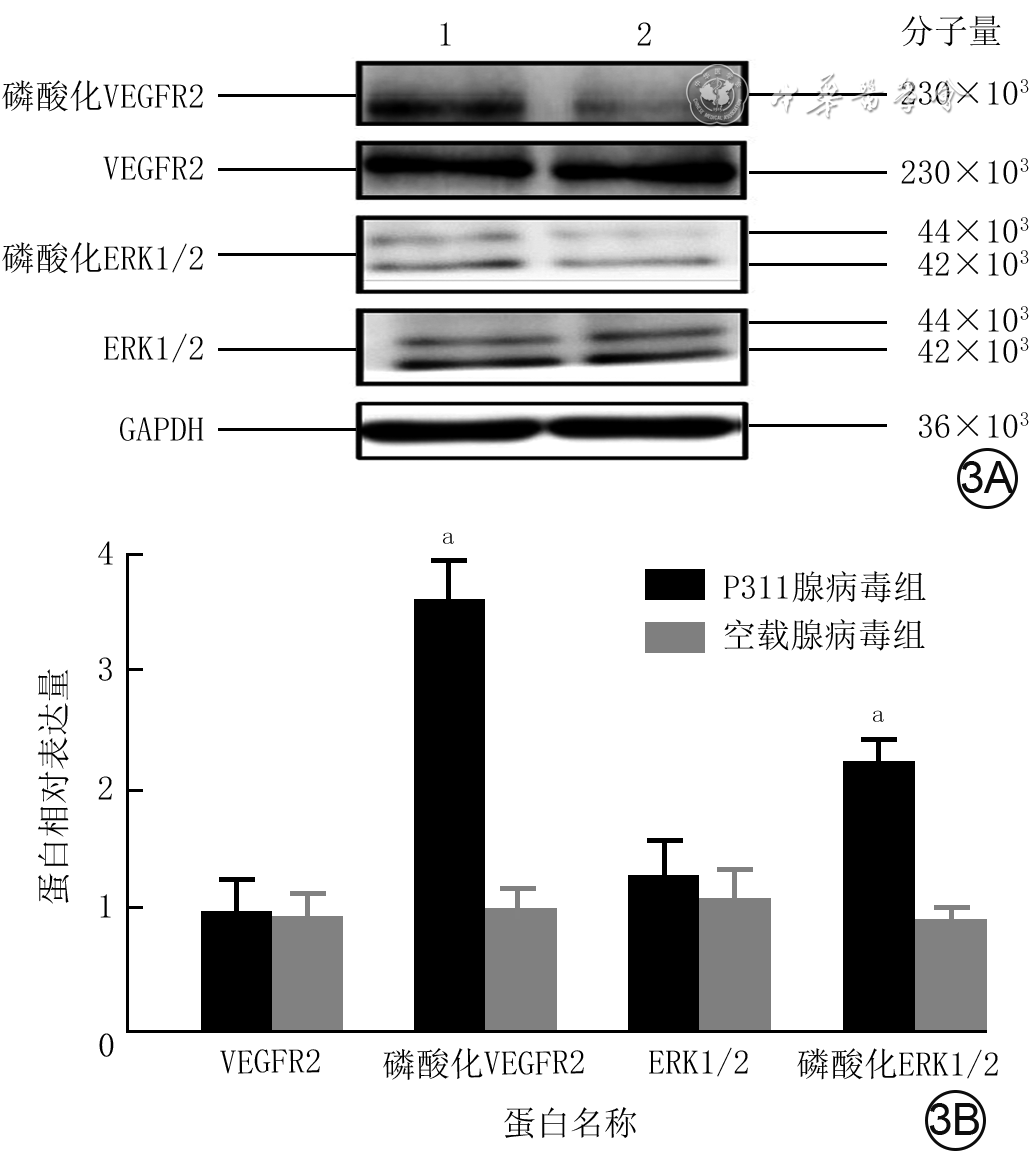
转染48 h,与空载腺病毒组比较,P311腺病毒组细胞VEGFR2、ERK1/2蛋白表达量均无明显变化(t值分别为0.28、1.36,P值分别为0.786、0.202),磷酸化VEGFR2、磷酸化ERK1/2蛋白表达量均明显升高(t值分别为17.27、16.08,P<0.001)。见图3。
 3 蛋白质印迹法检测2组人微血管内皮细胞1转染48 h VEGFR2/ERK1/2信号通路相关蛋白的表达量。3A.条带图;3B.条图(样本数为6,
3 蛋白质印迹法检测2组人微血管内皮细胞1转染48 h VEGFR2/ERK1/2信号通路相关蛋白的表达量。3A.条带图;3B.条图(样本数为6,2.2 P311通过VEGFR2/ERK1/2信号通路影响HMEC-1体外血管形成能力
2.2.1 siRNA-VEGFR2对P311高表达引起的VEGFR2/ERK1/2信号通路活化的影响
转染24 h,P311腺病毒+阴性对照siRNA组、空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组和空载腺病毒+siRNA-VEGFR2组细胞磷酸化VEGFR2、VEGFR2和磷酸化ERK1/2蛋白表达量总体比较,差异均有统计学意义(F值分别为27.89、25.07、14.20,P<0.001);4组间细胞ERK1/2蛋白表达量总体比较,差异无统计学意义(F=0.08,P=0.971)。
转染24 h,P311腺病毒+阴性对照siRNA组细胞磷酸化VEGFR2和磷酸化ERK1/2蛋白表达量均明显高于空载腺病毒+阴性对照siRNA组(P值分别为<0.001、0.001),2组细胞VEGFR2和ERK1/2蛋白表达量相近(P值分别为0.063、0.640)。P311腺病毒+阴性对照siRNA组细胞VEGFR2、磷酸化VEGFR2、磷酸化ERK1/2蛋白表达量均明显高于P311腺病毒+siRNA-VEGFR2组(P<0.001),2组细胞ERK1/2蛋白表达量相近(P=0.822)。空载腺病毒+阴性对照siRNA组细胞VEGFR2、磷酸化ERK1/2蛋白表达量均明显高于空载腺病毒+siRNA-VEGFR2组(P值分别为<0.001、0.049),该2组细胞磷酸化VEGFR2和ERK1/2蛋白表达量相近(P值分别为0.071、0.761)。转染24 h,P311腺病毒+siRNA-VEGFR2组和空载腺病毒+siRNA-VEGFR2组细胞VEGFR2、磷酸化VEGFR2、ERK1/2和磷酸化ERK1/2蛋白表达量均相近(P值分别为0.155、0.493、0.952、0.314)。见图4。
 4 蛋白质印迹法检测4组人微血管内皮细胞1转染24 h VEGFR2/ERK1/2信号通路相关蛋白的表达量。4A.条带图;4B.条图(样本数为6,
4 蛋白质印迹法检测4组人微血管内皮细胞1转染24 h VEGFR2/ERK1/2信号通路相关蛋白的表达量。4A.条带图;4B.条图(样本数为6,2.2.2 siRNA-VEGFR2对P311高表达介导的血管形成能力的影响
转染24 h,P311腺病毒+阴性对照siRNA组细胞管状结构节点数明显多于空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组(P值均<0.01),管状结构总长度明显长于空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组(P值均<0.01)。转染24 h,空载腺病毒+阴性对照siRNA组细胞管状结构节点数明显多于空载腺病毒+siRNA-VEGFR2组(P<0.01),管状结构总长度明显长于空载腺病毒+siRNA-VEGFR2组(P<0.01)。P311腺病毒+siRNA-VEGFR2组和空载腺病毒+siRNA-VEGFR2组细胞管状结构节点数和管状结构总长度相近(P>0.05)。见图5、表3。
 5 4组人微血管内皮细胞1转染24 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。5A.P311腺病毒+阴性对照小干扰RNA(siRNA)组管状结构节点数较多,管状结构总长度较长;5B.空载腺病毒+阴性对照siRNA组管状结构节点数明显少于图5A,管状结构总长度明显短于图5A;5C.P311腺病毒+siRNA-血管内皮生长因子受体2(VEGFR2)组管状结构节点数明显少于图5A,管状结构总长度明显短于图5A;5D.空载腺病毒+siRNA-VEGFR2组管状结构节点数明显少于图5B,管状结构总长度明显短于图5B,且均与图5C相近表3 4组人微血管内皮细胞1转染24 h后血管形成情况(
5 4组人微血管内皮细胞1转染24 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。5A.P311腺病毒+阴性对照小干扰RNA(siRNA)组管状结构节点数较多,管状结构总长度较长;5B.空载腺病毒+阴性对照siRNA组管状结构节点数明显少于图5A,管状结构总长度明显短于图5A;5C.P311腺病毒+siRNA-血管内皮生长因子受体2(VEGFR2)组管状结构节点数明显少于图5A,管状结构总长度明显短于图5A;5D.空载腺病毒+siRNA-VEGFR2组管状结构节点数明显少于图5B,管状结构总长度明显短于图5B,且均与图5C相近表3 4组人微血管内皮细胞1转染24 h后血管形成情况(组别 样本数 管状结构节点数(个) 管状结构总长度(μm) P311腺病毒+阴性对照siRNA组 6 720±62 21 241±1 139 空载腺病毒+阴性对照siRNA组 6 428±38 17 005±1 156 P311腺病毒+siRNA- VEGFR2组 6 364±57 13 494±2 465 空载腺病毒+siRNA- VEGFR2组 6 310±75 11 600±2 776 F值 57.07 26.32 P值 <0.001 <0.001 P1值 <0.001 0.002 P2值 <0.001 <0.001 P3值 0.002 <0.001 P4值 0.127 0.121 注:F值、P值为4组间各指标总体比较所得;P1值、P2值分别为P311腺病毒+阴性对照小干扰RNA(siRNA)组与空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-血管内皮生长因子受体2(VEGFR2)组比较所得,P3值、P4值分别为空载腺病毒+siRNA-VEGFR2组与空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组比较所得 2.2.3 ERK1/2抑制剂对P311高表达引起的ERK1/2信号通路活化的影响
处理2 h,P311腺病毒+DMSO组、空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组和空载腺病毒+ERK1/2抑制剂组细胞磷酸化ERK1/2蛋白表达量总体比较,差异有统计学意义(F=16.79,P<0.001);4组细胞ERK1/2 蛋白表达量总体比较,差异无统计学意义(F=0.99,P=0.419)。
P311腺病毒+DMSO组细胞磷酸化ERK1/2蛋白表达量明显高于空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组(P值均<0.001),P311腺病毒+DMSO组与空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组细胞ERK1/2蛋白表达量均相近(P值分别为0.806、0.467)。空载腺病毒+DMSO组细胞磷酸化ERK1/2蛋白表达量明显高于空载腺病毒+ERK1/2抑制剂组(P=0.030),2组细胞ERK1/2 蛋白表达量相近(P=0.140)。P311腺病毒+ERK1/2 抑制剂组和空载腺病毒+ERK1/2抑制剂组细胞磷酸化ERK1/2和ERK1/2蛋白表达量均相近(P值分别为0.591、0.373)。见图6。
 6 蛋白质印迹法检测4组人微血管内皮细胞1处理2 h ERK1/2信号通路相关蛋白的表达量。6A.条带图;6B.条图(样本数为6,
6 蛋白质印迹法检测4组人微血管内皮细胞1处理2 h ERK1/2信号通路相关蛋白的表达量。6A.条带图;6B.条图(样本数为6,2.2.4 ERK1/2抑制剂对P311高表达介导的促HMEC-1体外血管形成作用的影响
于处理2 h,P311腺病毒+DMSO组细胞管状结构节点数明显多于空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组(P值均<0.01),管状结构总长度明显长于空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组(P值均<0.01)。空载腺病毒+DMSO组管状结构节点数明显多于空载腺病毒+ERK1/2抑制剂组(P<0.01),管状结构总长度明显长于空载腺病毒+ERK1/2抑制剂组(P<0.01)。P311腺病毒+ERK1/2 抑制剂组和空载腺病毒+ERK1/2抑制剂组细胞管状结构节点数和总长度均相近(P>0.05)。见图7、表4。
 7 4组人微血管内皮细胞1处理2 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。7A.P311腺病毒+二甲基亚砜(DMSO)组管状结构节点数较多,管状结构总长度较长;7B.空载腺病毒+DMSO组管状结构节点数少于图7A,管状结构总长度短于图7A;7C.P311腺病毒+胞外信号调节激酶1/2(ERK1/2)抑制剂组管状结构节点数少于图7A,管状结构总长度短于图7A;7D.空载腺病毒+ERK1/2抑制剂组管状结构节点数和总长度与图7C相近表4 4组人微血管内皮细胞1处理2 h血管形成情况(
7 4组人微血管内皮细胞1处理2 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。7A.P311腺病毒+二甲基亚砜(DMSO)组管状结构节点数较多,管状结构总长度较长;7B.空载腺病毒+DMSO组管状结构节点数少于图7A,管状结构总长度短于图7A;7C.P311腺病毒+胞外信号调节激酶1/2(ERK1/2)抑制剂组管状结构节点数少于图7A,管状结构总长度短于图7A;7D.空载腺病毒+ERK1/2抑制剂组管状结构节点数和总长度与图7C相近表4 4组人微血管内皮细胞1处理2 h血管形成情况(组别 样本数 管状结构节点数(个) 管状结构总长度(μm) P311腺病毒+DMSO组 6 726±72 20 318±1 433 空载腺病毒+DMSO组 6 421±39 16 846±1 464 P311腺病毒+ERK1/2抑制剂组 6 365±41 15 114±1 950 空载腺病毒+ERK1/2抑制剂组 6 317±67 13 188±2 306 F值 63.49 16.53 P值 <0.001 <0.001 P1值 <0.001 0.004 P2值 <0.001 <0.001 P3值 0.005 0.002 P4值 0.162 0.083 注:F值、P值为4组间各指标总体比较所得;DMSO为二甲基亚砜,ERK1/2为胞外信号调节激酶1/2;P1值、P2值分别为P311腺病毒+DMSO组与空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组比较所得,P3值、P4值分别为空载腺病毒+ERK1/2抑制剂组与空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组比较所得 3. 讨论
创面愈合过程被划分为4个连续而又有重叠的阶段,包括凝血期、炎症期、增殖期和重构期[25]。创面一旦形成,就开始了局部新的血液供应系统的建立,该进程贯穿整个创面修复过程[2],但其机制仍不完全清楚。本课题组前期研究表明,皮肤微血管内皮细胞在静息状态下低表达P311,在炎症刺激条件下高表达P311,进而参与创面血管新生[22]。本研究进一步证实,P311能够通过激活VEGFR2/ERK1/2 信号通路,发挥促进HMEC-1血管形成的作用。
P311基因是一种损伤环境诱导型基因,即在静息状态下低表达或者不表达,当受到外界环境刺激时表达升高,从而发挥相关生物学功能[22, 23]。该基因能够编码一种种属间高度保守的细胞内多功能蛋白[10]。在创面愈合过程中,P311持续高表达,可增强表皮干细胞Rho A和Rac1蛋白的活性[21],并激活TGF-β1/Smad信号通路[23],从而发挥促进创面修复作用。近年来研究表明,P311表达于创面皮肤微血管内皮细胞,P311基因敲除导致皮肤创面愈合过程中的血管新生减弱,进而导致创面愈合延迟;P311基因敲除皮肤微血管内皮细胞迁移及血管形成能力均减弱[22]。这些研究结果提示P311可能通过影响微血管内皮细胞的迁移和血管形成能力参与血管新生。微血管内皮细胞具有正常有效的增殖能力在血管形成中也至关重要[26]。
本研究用P311腺病毒转染HMEC-1,CCK-8法检测结果显示,P311腺病毒组和空载腺病毒组HMEC-1增殖能力没有明显差异;划痕试验结果显示,P311腺病毒组HMEC-1迁移能力较空载腺病毒组明显增强;体外血管形成实验结果显示,P311腺病毒组HMEC-1血管形成能力较空载腺病毒组明显增强。以上结果表明P311高表达能够显著提高HMEC-1的迁移能力和体外血管形成能力,而对HMEC-1增殖能力没有明显影响。
VEGFR2信号通路在启动血管内皮细胞各项生物学过程中发挥重要作用[27]。VEGFR2活化是VEGFR2同二聚化或异二聚化导致酪氨酸激酶的活化和受体细胞内结构域中酪氨酸残基的自磷酸化[28, 29]。VEGFR2活化后启动血管内皮细胞内多种信号通路[30]。研究表明,在小鼠中VEGFR2 Tyr1173磷酸化(人类中Tyr1175磷酸化)对ERK1/2 激活至关重要[31]。ERK1/2信号通路与细胞的成活[32]、迁移[33]、血管形成[34]等生物学过程密切相关[35]。然而,此前尚鲜见有关P311对VEGFR2活化和ERK1/2活化的影响的研究。本研究采用蛋白质印迹法检测P311腺病毒组和空载腺病毒组HMEC-1中VEGFR2、磷酸化VEGFR2、ERK1/2和磷酸化ERK1/2的蛋白表达水平,结果显示,P311高表达对HMEC-1 VEGFR2、ERK1/2的蛋白表达没有明显影响,但是却能够促进VEGFR2、ERK1/2的磷酸化。根据上述结果可以得出,P311高表达可以激活VEGFR2/ERK1/2信号通路,该机制可能参与了P311促HMEC-1体外血管形成的作用。
为进一步探讨P311介导的促体外血管形成作用是否通过激活VEGFR2/ERK1/2信号通路而发挥作用,本研究分别采用siRNA-VEGFR2和ERK抑制剂SCH772984阻断VEGFR2/ERK1/2信号通路的活化,随后采用蛋白质印迹法检测细胞VEGFR2磷酸化和ERK1/2磷酸化水平的变化,采用体外血管形成实验检测细胞血管形成能力的变化。结果显示,P311腺病毒+阴性对照siRNA组细胞磷酸化VEGFR2和磷酸化ERK1/2蛋白表达水平均明显高于空载腺病毒+阴性对照siRNA组,提示P311高表达能够促进VEGFR2/ERK1/2信号通路活化。在阻断VEGFR2途径后,VEGFR2和ERK1/2磷酸化水平均明显降低,HMEC-1体外血管形成能力明显减弱,提示siRNA-VEGFR2能够抑制P311高表达引起的VEGFR2/ERK1/2信号通路活化,进而抑制P311高表达介导的促HMEC-1体外血管形成作用。另外,在抑制ERK1/2信号通路后,ERK1/2磷酸化水平均明显降低,HMEC-1体外血管形成能力明显减弱,提示ERK1/2抑制剂能够抑制P311高表达引起的ERK1/2信号通路活化,进而抑制P311高表达介导的促HMEC-1体外血管形成作用。由此推测,P311通过激活VEGFR2/ERK1/2信号通路发挥促进HMEC-1血管形成的作用。
综上所述,P311在创面血管新生中发挥重要作用,能够通过激活VEGFR2/ERK1/2信号通路发挥促进HMEC-1血管形成的作用。本研究结果丰富了创面愈合理论,为促进皮肤创面修复提供了新思路、新线索与新靶点。
·《Burns & Trauma》好文推荐·
严重烫伤后肠道菌群移位的分子机制:囊性纤维化穿膜传导调节蛋白的作用
王淞:实施研究、起草文章;李海胜:实施研究和分析数据;钱卫、张小容:采集数据和统计分析;贺伟峰:酝酿和设计实验;罗高兴:获取研究经费和对文章的知识性内容作批评性审阅所有作者均声明不存在利益冲突严重烫伤后肠道缺血缺氧会引起肠上皮屏障受损,发生肠道细菌移位(EBT),从而导致严重的并发症,如SIRS、脓毒症和MOF的发生。囊性纤维化穿膜传导调节蛋白(CFTR)可因肠上皮细胞缺氧而表达下调,进而影响紧密连接蛋白的正常结构与功能,而紧密连接蛋白正是维持肠道屏障的关键结构。陆军军医大学(第三军医大学)第一附属医院全军烧伤研究所陈婧老师团队联合上海交通大学第九人民医院整复外科章一新教授团队近期在《Burns & Trauma》发文《Molecular mechanism mediating enteric bacterial translocation after severe burn: the role of cystic fibrosis transmembrane conductance regulator》,使用人Caco-2细胞系建立体外缺氧损伤模型,并建立C57小鼠严重烫伤模型,研究严重烫伤对小鼠肠黏膜屏障、CFTR和紧密连接蛋白表达的影响,同时选取DF 508小鼠(F508del CFTR基因突变小鼠)为体内模型,进一步证明CFTR在维持小鼠正常肠道屏障功能中的作用。研究结果显示,在缺氧条件下,人Caco-2细胞中CFTR的表达显著降低;胞外信号调节激酶(ERK)和核因子κB信号被激活;炎症因子(TNF-α、IL-1β和IL-8)分泌增加;带状闭合蛋白1(ZO-1)、闭合蛋白和E-钙黏合素的表达下调;跨上皮电阻值降低;并导致ZO-1在细胞膜上分布不连续,排列不规则。同样地,敲除CFTR导致了相似的改变,且敲除CFTR引起的炎症因子的上调和紧密连接蛋白(ZO-1和闭合蛋白)的下调,可以通过抑制特定的ERK或核因子κB而逆转。与此同时,在严重烫伤小鼠的肠道中观察到大量的促炎症介质分泌和EBT,进一步支持了体外实验结果。与野生型小鼠相比,DF 508小鼠回肠中的TNF-α、IL-1β和IL-8浓度升高。此外,维生素D3被证明可以保护肠道上皮屏障免受缺氧损伤,机制可能与其上调CFTR表达,从而抑制ERK通路激活,减少促炎性细胞因子释放有关。此项研究结果表明,严重烫伤后,肠上皮细胞通过CFTR/ERK/促炎性细胞因子途径调节细胞间紧密连接蛋白的表达,进而调节肠道菌群移位,维生素D3可对严重烫伤后的肠黏膜屏障起到保护作用。杨云稀,编译自《Burns & Trauma》,2021,8:tkaa042;孙炳伟,审校 -
参考文献
(35) [1] FuX.Wound healing center establishment and new technology application in improving the wound healing quality in China[J/OL].Burns Trauma,2020,8:tkaa038[2021-12-10]. https://pubmed.ncbi.nlm.nih.gov/33134399/. DOI: 10.1093/burnst/tkaa038. [2] TonnesenMG,FengX,ClarkRA.Angiogenesis in wound healing[J].J Investig Dermatol Symp Proc,2000,5(1):40-46.DOI: 10.1046/j.1087-0024.2000.00014.x. [3] VeithAP,HendersonK,SpencerA,et al.Therapeutic strategies for enhancing angiogenesis in wound healing[J].Adv Drug Deliv Rev,2019,146:97-125.DOI: 10.1016/j.addr.2018.09.010. [4] FaheyE,DoyleSL.IL-1 family cytokine regulation of vascular permeability and angiogenesis[J].Front Immunol,2019,10:1426.DOI: 10.3389/fimmu.2019.01426. [5] LangmannT.Cytokine signaling as key regulator of pathological angiogenesis in the eye[J].EBioMedicine,2021,73:103662.DOI: 10.1016/j.ebiom.2021.103662. [6] VelnarT,GradisnikL.Tissue augmentation in wound healing: the role of endothelial and epithelial cells[J].Med Arch,2018,72(6):444-448.DOI: 10.5455/medarh.2018.72.444-448. [7] GonzalezAC,CostaTF,AndradeZA,et al.Wound healing- aliterature review[J].An Bras Dermatol,2016,91(5):614-620.DOI: 10.1590/abd1806-4841.20164741. [8] JohnsonKE,WilgusTA.Vascular endothelial growth factor and angiogenesis in the regulation of cutaneous wound repair[J].Adv Wound Care (New Rochelle),2014,3(10):647-661.DOI: 10.1089/wound.2013.0517. [9] GreavesNS,AshcroftKJ,BaguneidM,et al.Current understanding of molecular and cellular mechanisms in fibroplasia and angiogenesis during acute wound healing[J].J Dermatol Sci,2013,72(3):206-217.DOI: 10.1016/j.jdermsci.2013.07.008. [10] WeiZ,HanC,LiH,et al.Molecular mechanism of mesenchyme homeobox 1 in transforming growth factor β1-induced P311 gene transcription in fibrosis[J].Front Mol Biosci,2020,7:59.DOI: 10.3389/fmolb.2020.00059. [11] LagaresD.P311 in scar wars: myofibroblasts lost without transforming growth factor β translation[J].Am J Respir Cell Mol Biol,2019,60(2):139-140.DOI: 10.1165/rcmb.2018-0255ED. [12] YaoZ,YangS,HeW,et al.P311 promotes renal fibrosis via TGFβ1/Smad signaling[J].Sci Rep,2015,5:17032.DOI: 10.1038/srep17032. [13] DuanFF,BarronG,MelitonA,et al.P311 promotes lung fibrosis via stimulation of transforming growth factor-β1, -β2, and -β3 translation[J].Am J Respir Cell Mol Biol,2019,60(2):221-231.DOI: 10.1165/rcmb.2018-0028OC. [14] StradiotL,MannaertsI,van GrunsvenLA.P311, friend, or foe of tissue fibrosis?[J].Front Pharmacol,2018,9:1151.DOI: 10.3389/fphar.2018.01151. [15] TaylorGA,RodriguizRM,GreeneRI,et al.Behavioral characterization of P311 knockout mice[J].Genes Brain Behav,2008,7(7):786-795.DOI: 10.1111/j.1601-183X.2008.00420.x. [16] McDonoughWS,TranNL,BerensME.Regulation of glioma cell migration by serine-phosphorylated P311[J].Neoplasia,2005,7(9):862-872.DOI: 10.1593/neo.05190. [17] KattaK,SembajweLF,Kusche-GullbergM.Potential role for Ext1-dependent heparan sulfate in regulating P311 gene expression in A549 carcinoma cells[J].Biochim Biophys Acta Gen Subj,2018,1862(6):1472-1481.DOI: 10.1016/j.bbagen.2018.03.024. [18] BadriKR,YueM,CarreteroOA,et al.Blood pressure homeostasis is maintained by a P311-TGF-β axis[J].J Clin Invest,2013,123(10):4502-4512.DOI: 10.1172/JCI69884. [19] FujitaniM,YamagishiS,CheYH,et al.P311 accelerates nerve regeneration of the axotomized facial nerve[J].J Neurochem,2004,91(3):737-744.DOI: 10.1111/j.1471-4159.2004.02738.x. [20] ZhaoL,LeungJK,YamamotoH,et al.Identification of P311 as a potential gene regulating alveolar generation[J].Am J Respir Cell Mol Biol,2006,35(1):48-54.DOI: 10.1165/rcmb.2005-0475OC. [21] YaoZ,LiH,HeW,et al.P311 accelerates skin wound reepithelialization by promoting epidermal stem cell migration through RhoA and Rac1 activation[J].Stem Cells Dev,2017,26(6):451-460.DOI: 10.1089/scd.2016.0249. [22] WangS,ZhangX,QianW,et al.P311 deficiency leads to attenuated angiogenesis in cutaneous wound healing[J].Front Physiol,2017,8:1004.DOI: 10.3389/fphys.2017.01004. [23] LiH,YaoZ,HeW,et al.P311 induces the transdifferentiation of epidermal stem cells to myofibroblast-like cells by stimulating transforming growth factor β1 expression[J].Stem Cell Res Ther,2016,7(1):175.DOI: 10.1186/s13287-016-0421-1. [24] ZhouD,LiuT,WangS,et al.Effects of IL-1β and TNF-α on the expression of P311 in vascular endothelial cells and wound healing in mice[J].Front Physiol,2020,11:545008.DOI: 10.3389/fphys.2020.545008. [25] TottoliEM,DoratiR,GentaI,et al.Skin wound healing process and new emerging technologies for skin wound care and regeneration[J].Pharmaceutics,2020,12(8):735. DOI: 10.3390/pharmaceutics12080735. [26] 曹晓赞,谢挺,陆树良.糖化碱性成纤维细胞生长因子影响人真皮微血管内皮细胞增殖和血管化效应的受体途径[J].中华烧伤杂志,2021,37(1):17-24.DOI: 10.3760/cma.j.cn501120-20200916-00412. [27] AbhinandCS,RajuR,SoumyaSJ,et al.VEGF-A/VEGFR2 signaling network in endothelial cells relevant to angiogenesis[J].J Cell Commun Signal,2016,10(4):347-354.DOI: 10.1007/s12079-016-0352-8. [28] LemmonMA,SchlessingerJ.Cell signaling by receptor tyrosine kinases[J].Cell,2010,141(7):1117-1134.DOI: 10.1016/j.cell.2010.06.011. [29] ShibuyaM.VEGFR and type-V RTK activation and signaling[J].Cold Spring Harb Perspect Biol,2013,5(10):a009092.DOI: 10.1101/cshperspect.a009092. [30] SimonsM,GordonE,Claesson-WelshL.Mechanisms and regulation of endothelial VEGF receptor signalling[J].Nat Rev Mol Cell Biol,2016,17(10):611-625.DOI: 10.1038/nrm.2016.87. [31] DellingerMT,BrekkenRA.Phosphorylation of Akt and ERK1/2 is required for VEGF-A/VEGFR2-induced proliferation and migration of lymphatic endothelium[J].PLoS One,2011,6(12):e28947.DOI: 10.1371/journal.pone.0028947. [32] BakerA,WyattD,BocchettaM,et al.Notch-1-PTEN-ERK1/2 signaling axis promotes HER2+ breast cancer cell proliferation and stem cell survival[J].Oncogene,2018,37(33):4489-4504.DOI: 10.1038/s41388-018-0251-y. [33] ValantiEK, Dalakoura-KaragkouniK, FotakisP, et al. Reconstituted HDL-apoE3 promotes endothelial cell migration through ID1 and its downstream kinases ERK1/2, AKT and p38 MAPK[J]. Metabolism, 2022,127:154954. DOI: 10.1016/j.metabol.2021.154954. [34] TangH,HeY,LiL,et al.Exosomal MMP2 derived from mature osteoblasts promotes angiogenesis of endothelial cells via VEGF/Erk1/2 signaling pathway[J].Exp Cell Res,2019,383(2):111541.DOI: 10.1016/j.yexcr.2019.111541. [35] ShinM,BeaneTJ,QuillienA,et al.Vegfa signals through ERK to promote angiogenesis, but not artery differentiation[J].Development,2016,143(20):3796-3805.DOI: 10.1242/dev.137919. -

1 2组人微血管内皮细胞1划痕后各时间点剩余划痕面积 倒置相差显微镜×200,图中标尺为100 μm。1A、1B、1C.分别为P311腺病毒组划痕后0(即刻)、6、11 h,随着划痕后时间的延长,剩余划痕面积逐渐减少,至划痕后11 h划痕基本愈合;1D、1E、1F.分别为空载腺病毒组划痕后0、6、11 h,图1E、1F剩余划痕面积分别较图1B、1C明显增大
2 2组人微血管内皮细胞1培养8 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。2A. P311腺病毒组管状结构节点数较多,管状结构总长度较长;2B.空载腺病毒组管状结构节点数少于图2A,管状结构总长度短于图2A
3 蛋白质印迹法检测2组人微血管内皮细胞1转染48 h VEGFR2/ERK1/2信号通路相关蛋白的表达量。3A.条带图;3B.条图(样本数为6,
注:VEGFR2为血管内皮生长因子受体2,ERK1/2为胞外信号调节激酶1/2,GAPDH为3-磷酸甘油醛脱氢酶;条带图上方1、2分别为P311腺病毒组和空载腺病毒组;与空载腺病毒组比较,aP<0.01

4 蛋白质印迹法检测4组人微血管内皮细胞1转染24 h VEGFR2/ERK1/2信号通路相关蛋白的表达量。4A.条带图;4B.条图(样本数为6,
注:VEGFR2为血管内皮生长因子受体2,ERK1/2为胞外信号调节激酶1/2,GAPDH为3-磷酸甘油醛脱氢酶;条带图上方1、2、3、4分别为P311腺病毒+阴性对照小干扰RNA(siRNA)组、空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA- VEGFR2组和空载腺病毒+siRNA-VEGFR2组;与P311腺病毒+阴性对照siRNA组比较,aP<0.01;与空载腺病毒+阴性对照siRNA组比较,bP<0.01,cP<0.05
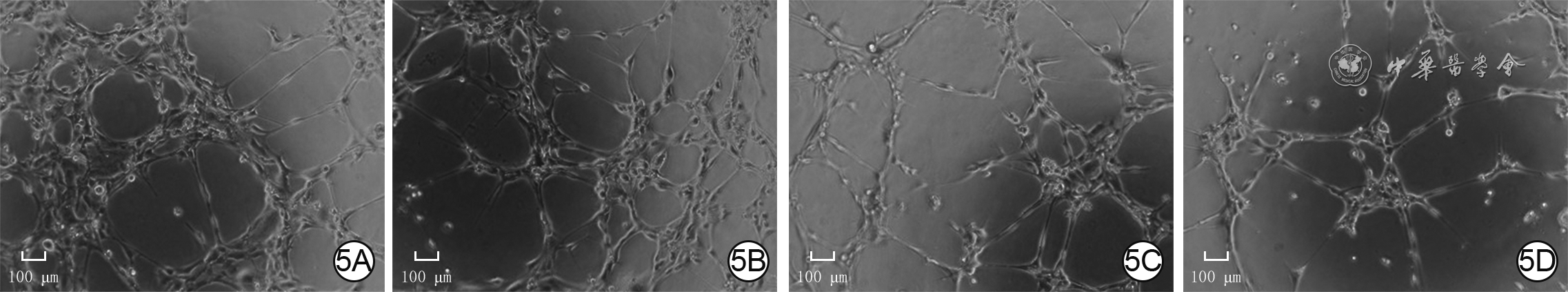
5 4组人微血管内皮细胞1转染24 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。5A.P311腺病毒+阴性对照小干扰RNA(siRNA)组管状结构节点数较多,管状结构总长度较长;5B.空载腺病毒+阴性对照siRNA组管状结构节点数明显少于图5A,管状结构总长度明显短于图5A;5C.P311腺病毒+siRNA-血管内皮生长因子受体2(VEGFR2)组管状结构节点数明显少于图5A,管状结构总长度明显短于图5A;5D.空载腺病毒+siRNA-VEGFR2组管状结构节点数明显少于图5B,管状结构总长度明显短于图5B,且均与图5C相近

6 蛋白质印迹法检测4组人微血管内皮细胞1处理2 h ERK1/2信号通路相关蛋白的表达量。6A.条带图;6B.条图(样本数为6,
注:F值、P值为4组间各指标总体比较所得;ERK1/2为胞外信号调节激酶1/2,GAPDH为3-磷酸甘油醛脱氢酶;条带图上方1、2、3、4分别为P311腺病毒+二甲基亚砜(DMSO)组、空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组和空载腺病毒+ERK1/2抑制剂组;与P311腺病毒+DMSO组比较,aP<0.01;与空载腺病毒+DMSO组比较,bP<0.05
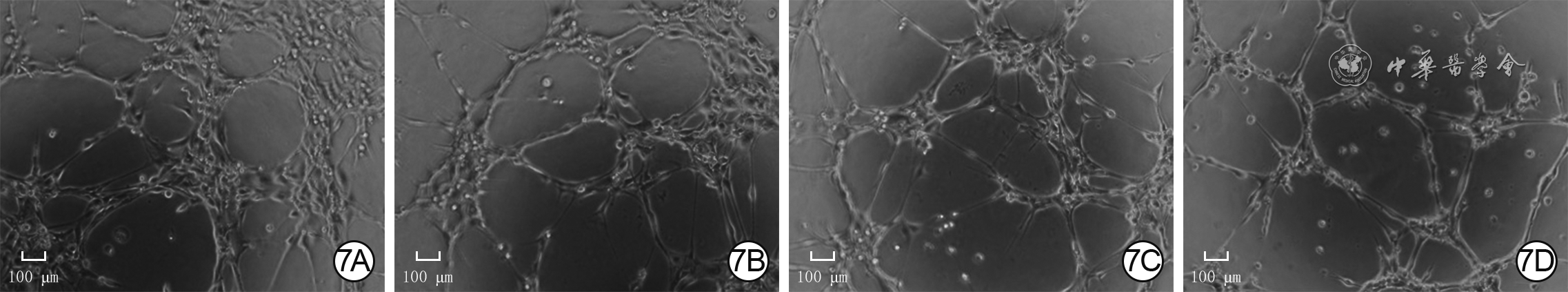
7 4组人微血管内皮细胞1处理2 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。7A.P311腺病毒+二甲基亚砜(DMSO)组管状结构节点数较多,管状结构总长度较长;7B.空载腺病毒+DMSO组管状结构节点数少于图7A,管状结构总长度短于图7A;7C.P311腺病毒+胞外信号调节激酶1/2(ERK1/2)抑制剂组管状结构节点数少于图7A,管状结构总长度短于图7A;7D.空载腺病毒+ERK1/2抑制剂组管状结构节点数和总长度与图7C相近
表1 2组人微血管内皮细胞1培养各时间点增殖活性比较(
组别 样本数 1 d 3 d 5 d P311腺病毒组 6 0.188±0.030 0.415±0.032 0.713±0.030 空载腺病毒组 6 0.185±0.019 0.392±0.030 0.687±0.031 t值 -0.23 -1.30 -1.52 P值 0.822 0.222 0.160 注:处理因素主效应,F=5.10,P=0.074;时间因素主效应,F=720.12,P<0.001;两者交互作用,F=0.81,P=0.473  下载: 导出CSV
下载: 导出CSV
表2 2组人微血管内皮细胞1划痕后各时间点剩余划痕面积百分比比较(%,
组别 样本数 6 h 11 h P311腺病毒组 6 47±7 9±5 空载腺病毒组 6 55±5 20±10 t值 -2.47 -2.62 P值 0.033 0.025 注:初始剩余划痕面积百分比均为100%;处理因素主效应,F=18.42,P=0.008;时间因素主效应,F=443.86,P<0.001;两者交互作用,F=7.89,P=0.041
下载: 导出CSV
表3 4组人微血管内皮细胞1转染24 h后血管形成情况(
组别 样本数 管状结构节点数(个) 管状结构总长度(μm) P311腺病毒+阴性对照siRNA组 6 720±62 21 241±1 139 空载腺病毒+阴性对照siRNA组 6 428±38 17 005±1 156 P311腺病毒+siRNA- VEGFR2组 6 364±57 13 494±2 465 空载腺病毒+siRNA- VEGFR2组 6 310±75 11 600±2 776 F值 57.07 26.32 P值 <0.001 <0.001 P1值 <0.001 0.002 P2值 <0.001 <0.001 P3值 0.002 <0.001 P4值 0.127 0.121 注:F值、P值为4组间各指标总体比较所得;P1值、P2值分别为P311腺病毒+阴性对照小干扰RNA(siRNA)组与空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-血管内皮生长因子受体2(VEGFR2)组比较所得,P3值、P4值分别为空载腺病毒+siRNA-VEGFR2组与空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组比较所得
下载: 导出CSV
表4 4组人微血管内皮细胞1处理2 h血管形成情况(
组别 样本数 管状结构节点数(个) 管状结构总长度(μm) P311腺病毒+DMSO组 6 726±72 20 318±1 433 空载腺病毒+DMSO组 6 421±39 16 846±1 464 P311腺病毒+ERK1/2抑制剂组 6 365±41 15 114±1 950 空载腺病毒+ERK1/2抑制剂组 6 317±67 13 188±2 306 F值 63.49 16.53 P值 <0.001 <0.001 P1值 <0.001 0.004 P2值 <0.001 <0.001 P3值 0.005 0.002 P4值 0.162 0.083 注:F值、P值为4组间各指标总体比较所得;DMSO为二甲基亚砜,ERK1/2为胞外信号调节激酶1/2;P1值、P2值分别为P311腺病毒+DMSO组与空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组比较所得,P3值、P4值分别为空载腺病毒+ERK1/2抑制剂组与空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组比较所得
下载: 导出CSV
-

 下载:
下载:
计量
- 文章访问数: 979
- HTML全文浏览量: 103
- PDF下载量: 32
- 被引次数: 0
