



Effects and mechanism of human umbilical vein endothelial cells-derived exosomes on wound healing in diabetic rabbits
-
摘要:
目的 探讨人脐静脉内皮细胞(HUVEC)外泌体对糖尿病兔创面愈合的作用及其机制。 方法 采用实验研究方法。取中南大学湘雅三医院2019年6月收治的2例糖尿病溃疡患者(男49岁、女58岁)手术切除溃疡周边皮肤组织,提取原代血管内皮细胞(VEC)和人皮肤成纤维细胞(HSF),通过形态观察和流式细胞术成功鉴定。采用超速离心法提取HUVEC外泌体,通过形态观察、粒径检测和蛋白质印迹法检测成功鉴定。取20只3个月龄雌性新西兰兔,背部两侧分别制作1个2型糖尿病全层皮肤缺损创面,将创面分成外泌体组和磷酸盐缓冲液(PBS)组并进行相应处理,每组20个创面,观察创面组织完全覆盖时间。伤后14 d,行苏木精-伊红染色或Masson染色,观察血管生成或胶原纤维增生情况(样本数为20)。观察VEC和HSF与HUVEC外泌体共培养24 h对HUVEC外泌体的摄取情况。将VEC与HSF均分成采用HUVEC外泌体或PBS处理的外泌体组和PBS组,采用细胞计数试剂盒8检测培养4 d细胞增殖情况,采用划痕试验检测并计算划痕后24、48 h细胞迁移率,采用Transwell实验检测培养24 h细胞迁移数,采用实时荧光定量反转录PCR法检测核因子红细胞系2相关因子2(NRF2)、转录激活因子3(ATF3)的mRNA表达,行成管实验观测VEC培养12 h血管分支点数和成管长度(样本数均为3)。取VEC及HSF,分成同前处理的PBS组、外泌体组和采用相应基因干扰的NRF2干扰组、ATF3干扰组、空载干扰组,同前检测2种细胞增殖、迁移和VEC的血管形成(样本数均为3)。对数据行重复测量方差分析、单因素方差分析、独立样本t检验及LSD检验。 结果 外泌体组创面组织完全覆盖时间为(17.9±1.9)d,明显短于PBS组的(25.2±2.3)d,t=4.54,P<0.05。伤后14 d,PBS组创面血管密度明显低于外泌体组(t=10.12,P<0.01),胶原纤维生成少于外泌体组。培养24 h,HUVEC外泌体成功被VEC和HSF摄取。培养4 d,外泌体组HSF和VEC增殖活力均明显强于PBS组(t值分别为54.73、7.05,P<0.01)。划痕后24、48 h,外泌体组HSF迁移率(t值分别为3.42、11.87,P<0.05或P<0.01)和VEC迁移率(t值分别为21.42、5.49,P<0.05或P<0.01)均明显高于PBS组。培养24 h,外泌体组VEC、HSF迁移数均明显多于PBS组(t值分别为12.31、16.78,P<0.01)。培养12 h,外泌体组HSF和VEC中NRF2的mRNA表达均明显高于PBS组(t值分别为7.52、5.78,P<0.05或P<0.01),ATF3的mRNA表达均明显低于PBS组(t值分别为13.44、8.99,P<0.01)。培养12 h,外泌体组VEC血管分支点数明显多于PBS组(t=17.60,P<0.01),成管长度明显长于PBS组(t=77.30,P<0.01)。培养4 d,NRF2干扰组HSF和VEC增殖活力均较PBS组和外泌体组显著降低(P<0.05或P<0.01);ATF3干扰组HSF和VEC增殖活力均较PBS组显著增加(P<0.05或P<0.01),均较外泌体组显著降低(P<0.05或P<0.01)。划痕后24、48 h,ATF3干扰组HSF和VEC的迁移率均较PBS组显著增加(P<0.05或P<0.01),均较外泌体组显著降低(P<0.05或P<0.01)。划痕后24、48 h,NRF2干扰组HSF和VEC的迁移率均较PBS组和外泌体组显著降低(P<0.05或P<0.01)。培养24 h,ATF3干扰组VEC和HSF的迁移数均明显多于PBS组(P<0.05),均明显少于外泌体组(P<0.05或P<0.01);NRF2干扰组VEC和HSF的迁移数均明显少于PBS组和外泌体组(P<0.01)。培养12 h,NRF2干扰组VEC血管长度、分支点数均较PBS组和外泌体组明显减少(P<0.01);ATF3干扰组VEC血管长度、分支点数均较PBS组明显增加(P<0.01),均较外泌体组明显减少(P<0.01)。 结论 HUVEC外泌体通过促进VEC和HSF的增殖、迁移,从而促进糖尿病兔创面愈合,而NRF2 和 ATF3 在这个过程中明显受到外泌体的影响,是外泌体作用的可能靶点。 Abstract:Objective The investigate the effects and mechanism of exosomes derived from human umbilical vein endothelial cells (HUVECs) on wound healing in diabetes rabbits. Methods The experimental research methods were used. The primary vascular endothelial cells (VECs) and human skin fibroblasts (HSFs) were extracted from skin tissue around ulcer by surgical excision of two patients with diabetic ulcer (the male aged 49 years and the female aged 58 years) admitted to Xiangya Third Hospital of Central South University in June 2019. The cells were successfully identified through morphological observation and flow cytometry. The HUVEC exosomes were extracted by ultracentrifugation and identified successfully by morphological observation, particle size detection, and Western blotting detection. Twenty female 3-month-old New Zealand rabbits were taken to create one type 2 diabetic full-thickness skin defect wound respectively on both sides of the back. The wounds were divided into exosomes group and phosphate buffer solution (PBS) group and treated accordingly, with 20 wounds in each group, the time of complete tissue coverage of wound was recorded. On PID 14, hematoxylin-eosin staining or Masson staining was performed to observe angiogenesis or collagen fiber hyperplasia (n=20). The VECs and HSFs were co-cultured with HUVEC exosomes for 24 h to observe the uptake of HUVEC exosomes by the two kinds of cells. The VECs and HSFs were divided to exosome group treated with HUVEC exosomes and PBS group treated with PBS to detect the cell proliferation on 4 d of culture with cell count kit 8, to detect and calculate the cell migration rate at 24 and 48 h after scratch by scratch test, to detect the cell migration number at 24 h of culture with Transwell test, and to detect the mRNA expressions of nuclear factor-erythroid 2-related factor 2 (NRF2) and transcription activating factor 3 (ATF3) by real time fluorescence quantitative reverse transcription polymerase chain reaction. Besides, the number of vascular branches and vascular length were observed in the tube forming experiment after 12 h of culture of VECs (n=3). The VECs and HSFs were taken and divided into PBS group and exosome group treated as before, and NRF2 interference group, ATF3 interference group, and no-load interference group with corresponding gene interference. The proliferation and migration of the two kinds of cells, and angiogenesis of VECs were detected as before (n=3). Data were statistically analyzed with analysis of variance for repeated measurement, one-way analysis of variance, independent sample t test, and least significant difference test. Results The time of complete tissue coverage of wound in exosome group was (17.9±1.9) d, which was significantly shorter than (25.2±2.3) d in PBS group (t=4.54, P<0.05). On PID14, the vascular density of wound in PBS group was significantly lower than that in exosome group (t=10.12, P<0.01), and the collagen fiber hyperplasia was less than that in exosome group. After 24 h of culture, HUVEC exosomes were successfully absorbed by VECs and HSFs. The proliferative activity of HSFs and VECs in exosome group was significantly higher than that in PBS group after 4 d of culture (with t values of 54.73 and 7.05, respectively, P<0.01). At 24 and 48 h after scratch, the migration rates of HSFs (with t values of 3.42 and 11.87, respectively, P<0.05 or P<0.01) and VECs (with t values of 21.42 and 5.49, respectively, P<0.05 or P<0.01) in exosome group were significantly higher than those in PBS group. After 24 h of culture, the migration numbers of VECs and HSFs in exosome group were significantly higher than those in PBS group (with t values of 12.31 and 16.78, respectively, P<0.01). After 12 h of culture, the mRNA expressions of NRF2 in HSFs and VECs in exosome group were significantly higher than those in PBS group (with t values of 7.52 and 5.78, respectively, P<0.05 or P<0.01), and the mRNA expressions of ATF3 were significantly lower than those in PBS group (with t values of 13.44 and 8.99, respectively, P<0.01). After 12 h of culture, the number of vascular branches of VECs in exosome group was significantly more than that in PBS group (t=17.60, P<0.01), and the vascular length was significantly longer than that in PBS group (t=77.30, P<0.01). After 4 d of culture, the proliferation activity of HSFs and VECs in NRF2 interference group was significantly lower than that in PBS group and exosome group (P<0.05 or P<0.01); the proliferation activity of HSFs and VECs in ATF3 interference group was significantly higher than that in PBS group (P<0.05 or P<0.01) and significantly lower than that in exosome group (P<0.05 or P<0.01). At 24 and 48 h after scratch, the migration rates of HSFs and VECs in ATF3 interference group were significantly higher than those in PBS group (P<0.05 or P<0.01) and significantly lower than those in exosome group (P<0.05 or P<0.01). At 24 and 48 h after scratch, the migration rates of HSFs and VECs in NRF2 interference group were significantly lower than those in PBS group and exosome group (P<0.05 or P<0.01). After 24 h of culture, the migration numbers of VECs and HSFs in ATF3 interference group were significantly more than those in PBS group (P<0.05) and significantly less than those in exosome group (P<0.05 or P<0.01); the migration numbers of VECs and HSFs in NRF2 interference group were significantly less than those in PBS group and exosome group (P<0.01). After 12 h of culture, the vascular length and number of branches of VECs in NRF2 interference group were significantly decreased compared with those in PBS group and exosome group (P<0.01); the vascular length and number of branches of VECs in ATF3 interference group were significantly increased compared with those in PBS group (P<0.01) and were significantly decreased compared with those in exosome group (P<0.01). Conclusions HUVEC exosomes can promote the wound healing of diabetic rabbits by promoting the proliferation and migration of VECs and HSFs, and NRF2 and ATF3 are obviously affected by exosomes in this process, which are the possible targets of exosome action. -
参考文献
(31) [1] MezeraV,BurešI.Chronic non-healing wounds in geriatrics[J].Vnitr Lek,2018,64(11):1098-1104. [2] HanG,CeilleyR.Chronic wound healing: a review of current management and treatments[J].Adv Ther,2017,34(3):599-610.DOI: 10.1007/s12325-017-0478-y. [3] GreeningDW,GopalSK,XuR,et al.Exosomes and their roles in immune regulation and cancer[J].Semin Cell Dev Biol,2015,40:72-81.DOI: 10.1016/j.semcdb.2015.02.009. [4] KalaniA,ChaturvediP,KamatPK,et al.Curcumin-loaded embryonic stem cell exosomes restored neurovascular unit following ischemia-reperfusion injury[J].Int J Biochem Cell Biol,2016,79:360-369.DOI: 10.1016/j.biocel.2016.09.002. [5] ShabbirA,CoxA,Rodriguez-MenocalL,et al.Mesenchymal stem cell exosomes induce proliferation and migration of normal and chronic wound fibroblasts, and enhance angiogenesis in vitro[J].Stem Cells Dev,2015,24(14):1635-1647.DOI: 10.1089/scd.2014.0316. [6] HeX,DongZ,CaoY,et al.MSC-derived exosome promotes M2 polarization and enhances cutaneous wound healing[J].Stem Cells Int,2019,2019:7132708.DOI: 10.1155/2019/7132708. [7] CooperDR,WangC,PatelR,et al.Human adipose-derived stem cell conditioned media and exosomes containing MALAT1 promote human dermal fibroblast migration and ischemic wound healing[J].Adv Wound Care (New Rochelle),2018,7(9):299-308.DOI: 10.1089/wound.2017.0775. [8] ShafeiS,KhanmohammadiM,HeidariR,et al.Exosome loaded alginate hydrogel promotes tissue regeneration in full-thickness skin wounds: an in vivo study[J].J Biomed Mater Res A,2020,108(3):545-556.DOI: 10.1002/jbm.a.36835. [9] WangC,LiangC,WangR,et al.The fabrication of a highly efficient self-healing hydrogel from natural biopolymers loaded with exosomes for the synergistic promotion of severe wound healing[J].Biomater Sci,2019,8(1):313-324.DOI: 10.1039/c9bm01207a. [10] YousefiS,LiuT,WangRK.Segmentation and quantification of blood vessels for OCT-based micro-angiograms using hybrid shape/intensity compounding[J].Microvasc Res,2015,97:37-46.DOI: 10.1016/j.mvr.2014.09.007. [11] Romana-SouzaB,Assis de BritoTL,PereiraGR,et al.Gonadal hormones differently modulate cutaneous wound healing of chronically stressed mice[J].Brain Behav Immun,2014,36:101-110.DOI: 10.1016/j.bbi.2013.10.015. [12] BjørgeIM,KimSY,ManoJF,et al.Extracellular vesicles, exosomes and shedding vesicles in regenerative medicine - a new paradigm for tissue repair[J].Biomater Sci,2017,6(1):60-78.DOI: 10.1039/c7bm00479f. [13] KatagiriS,ParkK,MaedaY,et al.Overexpressing IRS1 in endothelial cells enhances angioblast differentiation and wound healing in diabetes and insulin resistance[J].Diabetes,2016,65(9):2760-2771.DOI: 10.2337/db15-1721. [14] MurakamiS,MotohashiH.Roles of Nrf2 in cell proliferation and differentiation[J].Free Radic Biol Med,2015,88(Pt B):168-178.DOI: 10.1016/j.freeradbiomed.2015.06.030. [15] YiT,LiX,WangE,et al.Activation of the nuclear erythroid 2-related factor 2 antioxidant responsive element (Nrf2-ARE) signaling pathway alleviates acute graft-versus-host disease by reducing oxidative stress and inhibiting infiltration of inflammatory cells in an allogeneic stem cell transplantation mouse model[J].Med Sci Monit,2018,24:5973-5979.DOI: 10.12659/MSM.908130. [16] TonelliC,ChioIIC,TuvesonDA.Transcriptional regulation by Nrf2[J].Antioxid Redox Signal,2018,29(17):1727-1745.DOI: 10.1089/ars.2017.7342. [17] AmbrozovaN,UlrichovaJ,GalandakovaA.Models for the study of skin wound healing. The role of Nrf2 and NF-κB[J].Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub,2017,161(1):1-13.DOI: 10.5507/bp.2016.063. [18] LabzinLI,SchmidtSV,MastersSL,et al.ATF3 is a key regulator of macrophage IFN responses[J].J Immunol,2015,195(9):4446-4455.DOI: 10.4049/jimmunol.1500204. [19] ShaH,ZhangD,ZhangY,et al.ATF3 promotes migration and M1/M2 polarization of macrophages by activating tenascin-C via Wnt/β-catenin pathway[J].Mol Med Rep,2017,16(3):3641-3647.DOI: 10.3892/mmr.2017.6992. [20] HuY,RaoSS,WangZX,et al.Exosomes from human umbilical cord blood accelerate cutaneous wound healing through miR-21-3p-mediated promotion of angiogenesis and fibroblast function[J].Theranostics,2018,8(1):169-184.DOI: 10.7150/thno.21234. [21] MichalikKM,YouX,ManavskiY,et al.Long noncoding RNA MALAT1 regulates endothelial cell function and vessel growth[J].Circ Res,2014,114(9):1389-1397.DOI: 10.1161/CIRCRESAHA.114.303265. [22] AlamandaV,SinghS,LawrenceNJ,et al.Nicotine-mediated induction of E-selectin in aortic endothelial cells requires Src kinase and E2F1 transcriptional activity[J].Biochem Biophys Res Commun,2012,418(1):56-61.DOI: 10.1016/j.bbrc.2011.12.127. [23] WuX,ShowiheenS,SunAR,et al.Exosomes extraction and identification[J].Methods Mol Biol,2019,2054:81-91.DOI: 10.1007/978-1-4939-9769-5_4. [24] TongJ,YangF,LiX,et al.Mechanical characterization and material modeling of diabetic aortas in a rabbit model[J].Ann Biomed Eng,2018,46(3):429-442.DOI: 10.1007/s10439-017-1955-9. [25] SedovaKA,AzarovJE,ArteyevaNV,et al.Mechanism of electrocardiographic T-wave flattening in diabetes mellitus: experimental and simulation study[J].Physiol Res,2017,66(5):781-789.DOI: 10.33549/physiolres.933494. [26] ZhangY,BiJ,HuangJ,et al.Exosome: a review of its classification, isolation techniques, storage, diagnostic and targeted therapy applications[J].Int J Nanomedicine,2020,15:6917-6934.DOI: 10.2147/IJN.S264498. [27] WeiP, ZhongC, YangX, et al. Exosomes derived from human amniotic epithelial cells accelerate diabetic wound healing via PI3K-AKT-mTOR-mediated promotion in angiogenesis and fibroblast function[J/OL]. Burns Trauma, 2020,8:tkaa020[2022-10-31]. https://pubmed.ncbi.nlm.nih.gov/32923490/.DOI: 10.1093/burnst/tkaa020. [28] 卫裴, 许钊荣, 陈艺敏, 等. 人羊膜上皮细胞外泌体对高糖环境下HaCaT增殖和迁移的影响及其机制 [J] . 中华烧伤杂志, 2021, 37(12) : 1175-1184. DOI: 10.3760/cma.j.cn501120-20210424-00154. [29] TangPM,ZhouS,MengXM,et al.Smad3 promotes cancer progression by inhibiting E4BP4-mediated NK cell development[J].Nat Commun,2017,8:14677.DOI: 10.1038/ncomms14677. [30] GonçalvesLR,de Oliveira da Silva B,ScarinciLD,et al.MicroRNA-1254 contributes to the controlling of pro-fibrogenic environment in LX-2 cells by modulating SMAD3 and wound repair: new insights in hepatic fibrosis[J].Cell Biol Int,2019,43(3):333-343.DOI: 10.1002/cbin.11075. [31] ChenC, GeC, LiuZ,et al. ATF3 inhibits the tumorigenesis and progression of hepatocellular carcinoma cells via upregulation of CYR61 expression[J]. J Exp Clin Cancer Res,2018,37(1):263. DOI: 10.1186/s13046-018-0919-8. -

1 从糖尿病足溃疡患者创面周围组织提取的原代血管内皮细胞(VEC)和人皮肤成纤维细胞(HSF)形态 光学显微镜×400。1A.原代VEC呈菱形或多角形;1B.原代HSF呈纺锤形或者多角形扁平状

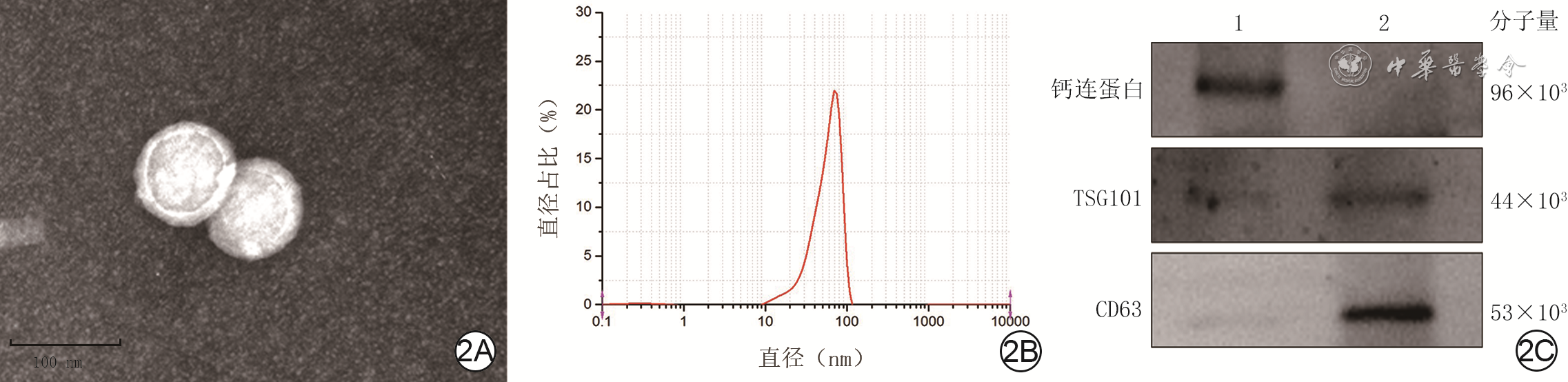
2 采用3种方法进行人脐静脉内皮细胞(HUVEC)外泌体的鉴定。2A.颗粒呈囊状或球形 电子显微镜×100 000;2B.粒度分析显示颗粒直径为70~100 nm;2C.蛋白质印迹法检测显示,较于人胚肾细胞293,HUVEC来源的颗粒高表达CD63和TSG101,低表达钙连蛋白
注:图2C为横坐标经过lg处理的数据形成的描记图,该图上方1、2分别为人胚肾细胞293与HUVEC来源外泌体;TSG101为肿瘤易感基因101

3 外泌体组和磷酸盐缓冲液(PBS)组全层皮肤缺损兔伤后各时间点创面愈合情况。3A、3B、3C.分别为伤后0(即刻)、14、30 d创面愈合情况,每张图中左侧标记A的为外泌体组,右侧标记B的为PBS组,外泌体组创面伤后14、30 d均较PBS组愈合快

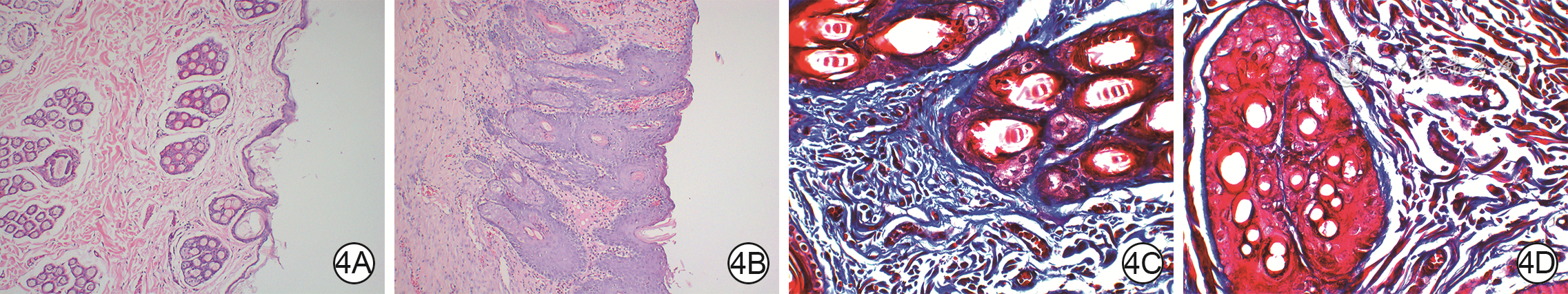
4 外泌体组和磷酸盐缓冲液(PBS)组全层皮肤缺损兔伤后14 d血管新生和胶原纤维增生情况。4A、4B.分别为外泌体组和PBS组血管新生情况 苏木精-伊红×200,图4A新生血管较图4B多;4C、4D.分别为外泌体组和PBS组胶原纤维增生情况 Masson×200,图4C胶原纤维增生较图4D多

5 从糖尿病足溃疡患者创面周围组织提取的血管内皮细胞(VEC)和人皮肤成纤维细胞(HSF)培养24 h成功摄取人脐静脉内皮细胞(HUVEC)外泌体 PKH67-鬼笔环肽-4',6-二脒基-2-苯基吲哚×200。5A、5B.分别为VEC及HSF胞质染色(红色)、胞核染色(蓝色)、外泌体染色(绿色)合并图,2种细胞均成功摄取外泌体

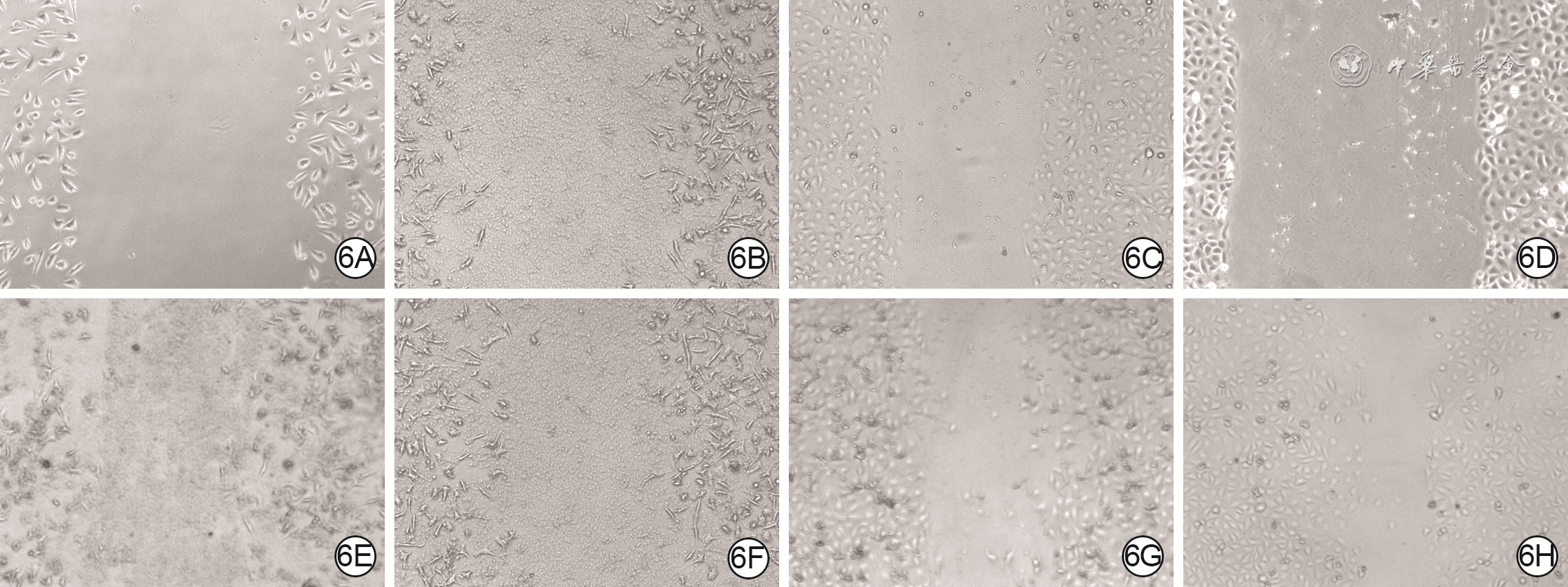
6 划痕试验观察2组人皮肤成纤维细胞(HSF)和血管内皮细胞(VEC)培养各时间点划痕情况 光学显微镜×200。6A、6B.分别为磷酸盐缓冲液(PBS)组和外泌体组HSF培养0 h(即刻);6C、6D.分别为PBS组和外泌体VEC培养0 h;6E、6F和6G、6H.分别为PBS组、外泌体组HSF和VEC培养48 h,外泌体组2种细胞培养48 h剩余划痕面积均小于PBS组

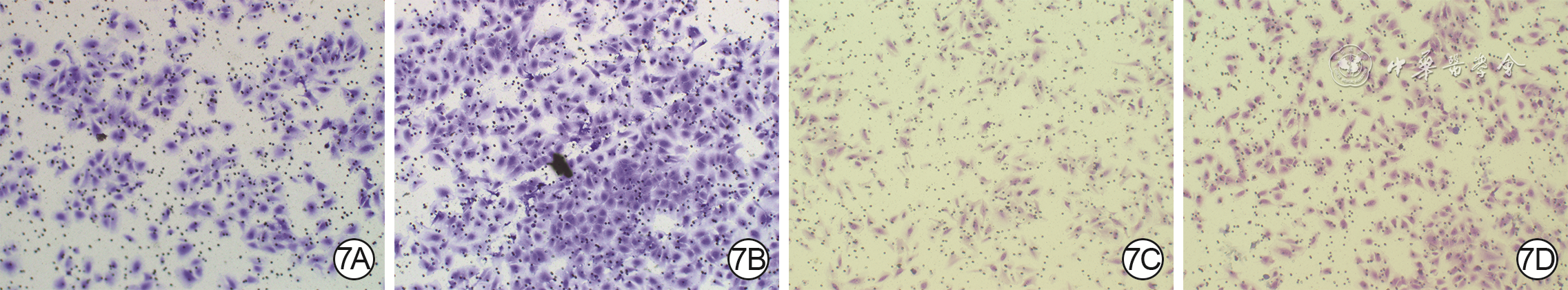
7 Transwell实验观察2组血管内皮细胞(VEC)和人皮肤成纤维细胞(HSF)培养24 h的迁移情况 结晶紫×200。7A.磷酸盐缓冲液(PBS)组VEC;7B.外泌体组VEC,迁移数明显多于图7A;7C.PBS组HSF;7D.外泌体组HSF,穿膜数明显多于图7C

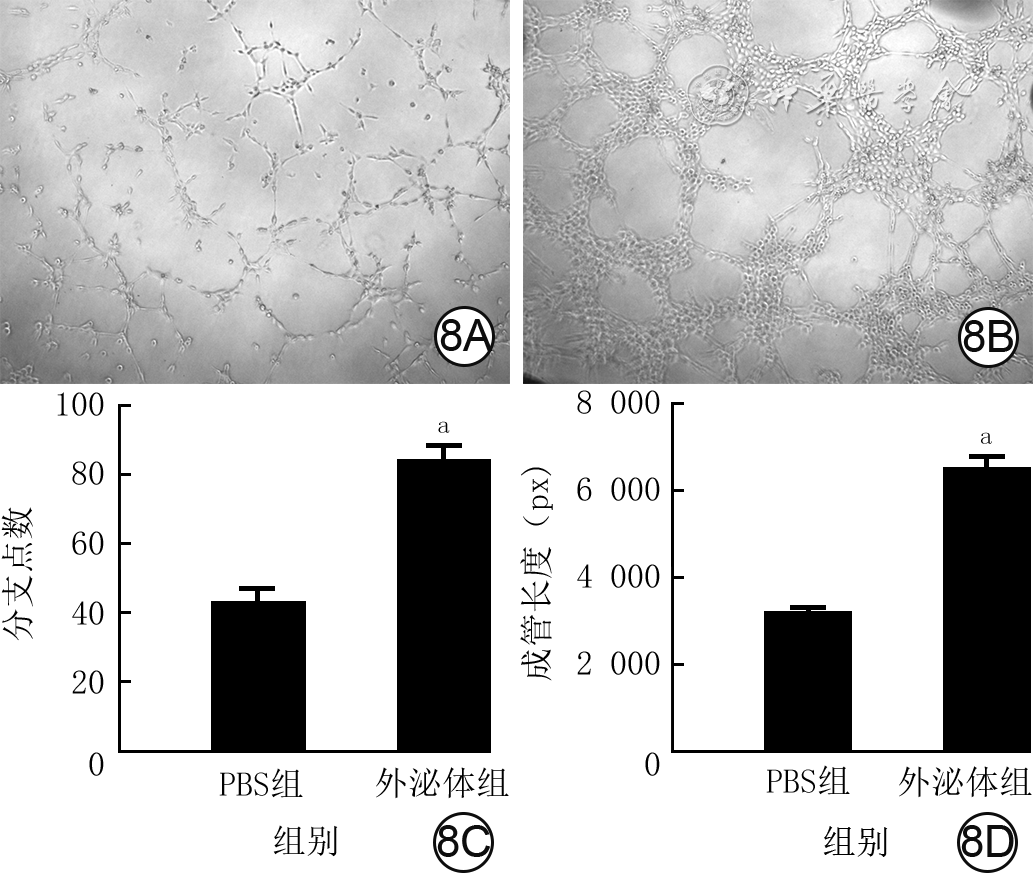
8 2组血管内皮细胞培养12 h血管分支点数和成管长度。8A、8B.分别为磷酸盐缓冲液(PBS)组和外泌体组血管生成情况,图8B新生血管数明显多于图8A 光学显微镜×400;8C、8D.分别为2组血管分支点数和成管长度比较(样本数为3,
注:与PBS组比较,aP<0.01

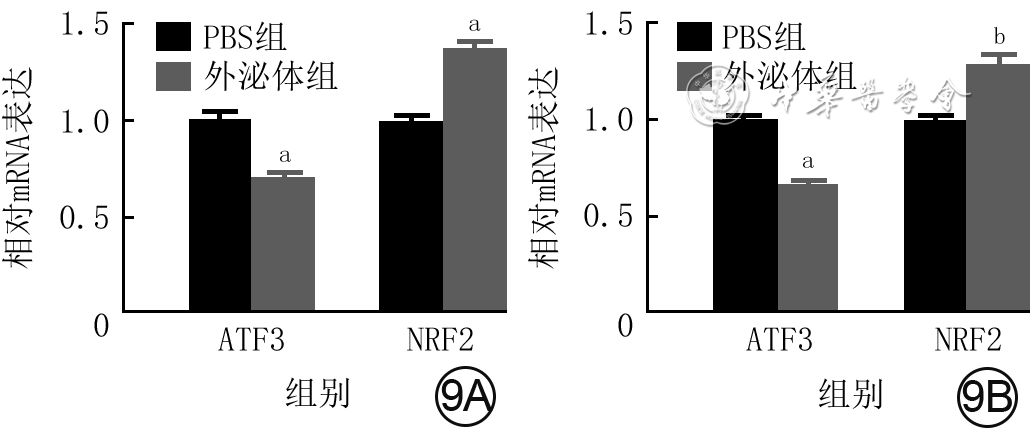
9 2组人皮肤成纤维细胞(HSF)和血管内皮细胞(VEC)培养12 h的核转录因子红系2相关因子2 (NRF2) 和转录激活因子3 (ATF3) mRNA表达比较(样本数为3,
注:与磷酸盐缓冲液(PBS)组比较,aP<0.01,bP<0.05

10 划痕试验观察5组人皮肤成纤维细胞(HSF)和血管内皮细胞(VEC)划痕后各时间点迁移情况 光学显微镜×200。10A、10B、10C、10D、10E及10F、10G、10H、10I、10J.分别为磷酸盐缓冲液(PBS)组、外泌体组、核转录因子红系2相关因子2 (NRF2) 干扰组、转录激活因子3 (ATF3) 干扰组、空载干扰组HSF划痕后0(即刻)、48 h;10K、10L、10M、10N、10O及10P、10Q、10R、10S、10T.分别为PBS组、外泌体组、NRF2干扰组、ATF3干扰组、空载干扰组VEC划痕后0、48 h

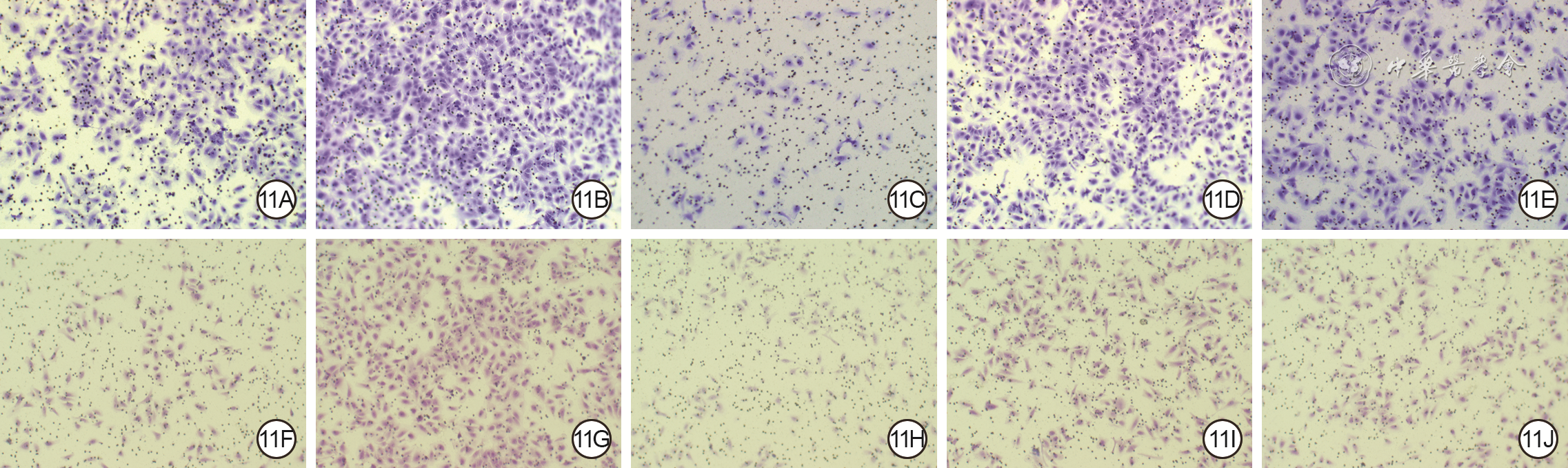
11 Transwell实验观察5组血管内皮细胞(VEC)和人皮肤成纤维细胞(HSF)培养24 h的迁移情况 结晶紫×200。11A、11B、11C、11D、11E.分别为磷酸盐缓冲液(PBS)组、外泌体组、核转录因子红系2相关因子2 (NRF2) 干扰组、转录激活因子3 (ATF3) 干扰组、空载干扰组VEC,图11C迁移数少于图11A和11B,图11D迁移数多于图11A、少于11B;11F、11G、11H、11I、11J.分别为PBS组、外泌体组、NRF2干扰组、ATF3干扰组、空载干扰组HSF,图11H迁移数少于图11F和11G,图11I迁移数多于图11F、少于图11G

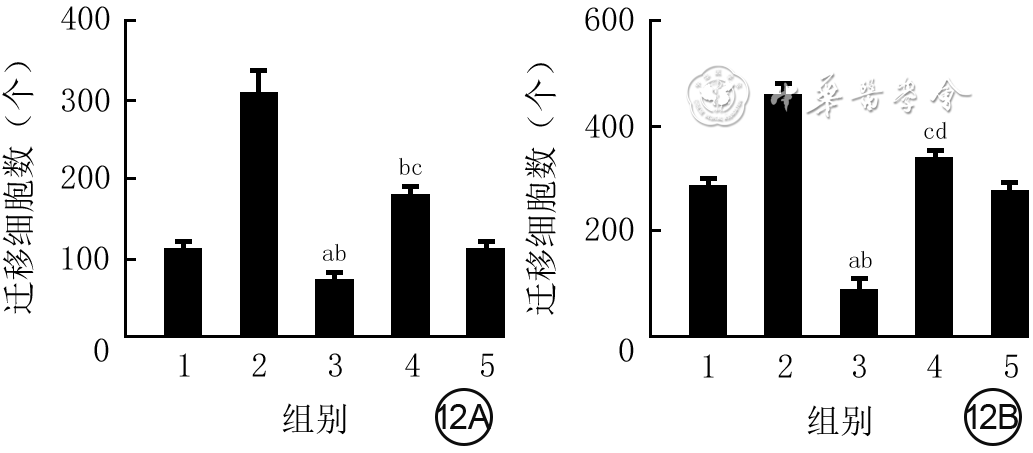
12 5组VEC和HSF培养24 h的迁移数比较(样本数为3,
注:横坐标下1、2、3、4、5分别代表磷酸盐缓冲液(PBS)组、外泌体组、核转录因子红系2相关因子2(NRF2)干扰组、转录激活因子3(ATF3)干扰组、空载干扰组;5组血管内皮细胞(VEC)和人皮肤成纤维细胞(HSF)总体比较,差异均明显(F值分别为11.43、21.10,P值分别为0.008、<0.001);与PBS组比较,aP<0.01,cP<0.05;与外泌体组比较,bP<0.01,dP<0.05

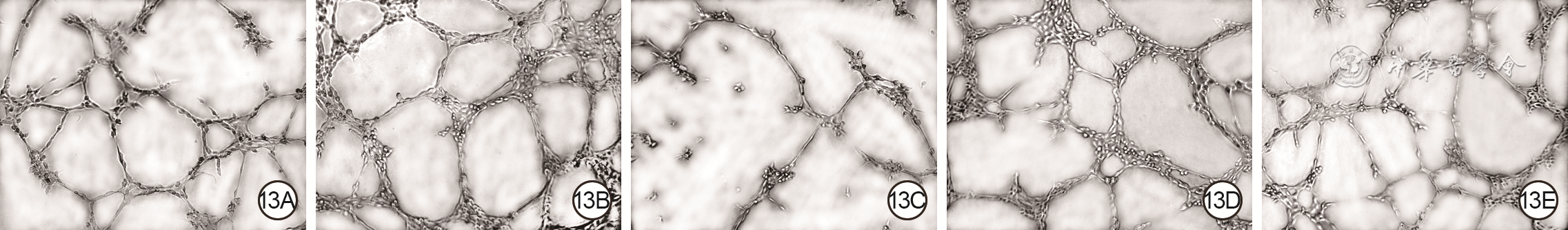
13 5组血管内皮细胞培养12 h血管分支点数和成管长度 光学显微镜×400。13A、13B、13C、13D、13E.分别为磷酸盐缓冲液组、外泌体组、核因子红细胞系2相关因子2干扰组、转录激活因子干扰组、空载干扰组,图13C成管长度和分支点数均较图13A、13B减少,图13D成管长度和分支点数均较图13A增加,较图13B减少

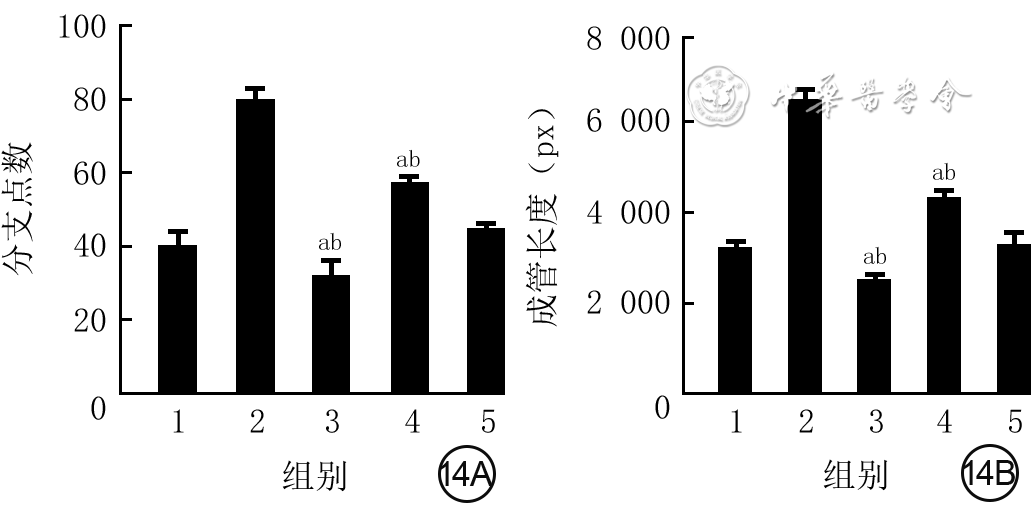
14 5组血管内皮细胞血管长度及分支数目比较(样本数为3,
注:横坐标下1、2、3、4、5分别代表磷酸盐缓冲液(PBS)组、外泌体组、核转录因子红系2相关因子2干扰组、转录激活因子3 干扰组、空载干扰组;5组血管分支点数和成管长度总体比较差异均明显(F值分别为7.51、9.12,P值分别为0.011、0.005);与PBS组比较,aP<0.01,与外泌体组比较,bP<0.01
表1 2组VEC和HSF划痕后各时间点迁移率比较(%,
组别 样本数 HSF VEC 24 h 48 h 24 h 48 h 外泌体组 3 42.1±1.3a 72.2±0.9b 48.2±1.6b 87.1±2.3b PBS组 3 26.1±1.3 44.2±1.0 30.1±1.6 63.2±1.5 注:人皮肤成纤维细胞(HSF)时间因素主效应,F=33.20,P=0.020;处理因素主效应,F=134.80,P=0.012;两者交互作用,F=95.10,P=0.021;血管内皮细胞(VEC)时间因素主效应,F=37.10,P=0.010;处理因素主效应,F=191.70,P=0.007;两者交互作用,F=45.10,P=0.009;与磷酸盐缓冲液(PBS)组比较,aP<0.05,bP<0.01  下载: 导出CSV
下载: 导出CSV
表2 5组血管内皮细胞和人皮肤成纤维细胞培养4 d增殖活力比较(
组别 样本数 人皮肤成纤维细胞 血管内皮细胞 PBS组 3 0.904±0.005 1.047±0.046 外泌体组 3 1.205±0.011 1.311±0.010 NRF2干扰组 3 0.813±0.006ab 0.896±0.005ad ATF3干扰组 3 1.003±0.005bc 1.192±0.014ad 空载干扰组 3 0.915±0.004 1.072±0.045 F值 7.98 6.02 P值 0.014 0.004 注:NRF2为核转录因子红系2相关因子2,ATF3为转录激活因子3;与磷酸盐缓冲液(PBS)组比较,aP<0.05,cP<0.01;与外泌体组比较,bP<0.01,dP<0.05
下载: 导出CSV
表3 5组血管内皮细胞和人皮肤成纤维细胞划痕后各时间点迁移率比较(%,
组别 样本数 人皮肤成纤维细胞 血管内皮细胞 24 h 48 h 24 h 48 h PBS组 3 25.2±0.9 41.2±1.0 31.2±2.0 61.2±1.0 外泌体组 3 40.8±1.1 69.2±0.7 49.8±1.0 86.0±2.0 NRF2干扰组 3 21.0±0.8ab 37.0±1.2bd 26.0±0.9ab 54.0±1.2bd ATF3干扰组 3 33.0±0.7ac 56.0±1.2cd 38.0±0.7ab 71.0±1.3bd 空载干扰组 3 25.0±0.7 40.0±1.1 30.0±1.0 60.0±1.0 注:NRF2为核转录因子红系2相关因子2,ATF3为转录激活因子3;人皮肤成纤维细胞时间因素主效应,F=33.20,P=0.022,处理因素主效应,F=104.60,P=0.003,两者交互作用,F=125.10,P=0.019;血管内皮细胞时间因素主效应,F=19.10,P=0.036,处理因素主效应,F=221.10,P=0.005,两者交互作用,F=45.10,P=0.009;与磷酸盐缓冲液(PBS)组比较,aP<0.05,dP<0.01;与外泌体组比较,bP<0.01,cP<0.05
下载: 导出CSV
-

 下载:
下载:






计量
- 文章访问数: 1447
- HTML全文浏览量: 170
- PDF下载量: 51
- 被引次数: 0
