



Effects and mechanism of glycine on rat cardiomyocytes pretreated with serum from burned rats
-
摘要:
目的 探讨甘氨酸对经烧伤大鼠血清(以下简称烧伤血清)干预的大鼠心肌细胞的作用及其机制。 方法 采用实验研究方法。取30只7~8周龄雌雄各半Wistar大鼠,其中10只用于制备正常大鼠血清(以下简称正常血清),将另20只造成30%体表总面积Ⅲ度烧伤后制备烧伤血清;从180只1~3 d龄雌雄不拘Wistar大鼠心尖组织中分离培养原代心肌细胞用于后续实验。按随机数字表法(分组方法下同)将细胞分为用相应血清处理的正常血清组、烧伤血清组,于处理1、3、6、9、12 h进行锥虫蓝染色检测细胞存活率;将细胞分为用烧伤血清处理6 h后常规培养30 min的单纯烧伤血清组和用烧伤血清处理6 h后加相应终物质的量浓度甘氨酸培养30 min的0.4 mmol/L甘氨酸组、0.8 mmol/L甘氨酸组、1.2 mmol/L甘氨酸组、1.6 mmol/L甘氨酸组、2.0 mmol/L甘氨酸组,即干预6.5 h,同前检测细胞存活率。将细胞分为正常血清组、单纯烧伤血清组、0.8 mmol/L甘氨酸组、1.2 mmol/L甘氨酸组、1.6 mmol/L甘氨酸组,分别同前干预6.5 h,采用高效液相色谱法检测腺苷一磷酸(AMP)和ATP含量并计算AMP/ATP比值,采用蛋白质印迹法检测磷酸化哺乳动物雷帕霉素靶蛋白复合物1(p-mTORC1)、磷酸化p70核糖体蛋白S6激酶(p-p70 S6K)、磷酸化真核翻译起始因子4E结合蛋白1(p-4E-BP1)、磷酸化AMP活化蛋白激酶(p-AMPK)蛋白表达。将细胞分为同前干预的正常血清组、单纯烧伤血清组、0.8 mmol/L甘氨酸组和用烧伤血清处理后加2种试剂培养的0.8 mmol/L甘氨酸+25 ng/mL雷帕霉素组,于处理1、3、6 h行相应培养30 min,即干预1.5、3.5、6.5 h,采用免疫荧光法检测热休克蛋白70(HSP70)、金属硫蛋白(MT)和微管蛋白表达并观察干预6.5 h微管形态。各时间点样本数均为10。对数据行析因设计方差分析、单因素方差分析、LSD- t检验、LSD检验及Bonferroni校正。 结果 处理1、3、6、9、12 h,烧伤血清组细胞存活率均明显低于正常血清组( t值分别为4.96、16.83、35.51、34.33、27.88, P<0.05)。在烧伤血清组中,处理3、6、9、12 h细胞存活率均较处理1 h明显降低( P<0.05),处理6、9、12 h细胞存活率均较处理3 h明显降低( P<0.05),处理6 h细胞存活率与处理9 h相近( P>0.05)但明显高于处理12 h( P<0.05);选择处理6 h作为后续烧伤血清干预时间。干预6.5 h,与单纯烧伤血清组比较,各甘氨酸组细胞存活率均明显升高( P<0.05)。0.8 mmol/L甘氨酸组细胞存活率最高,选择0.8、1.2、1.6 mmol/L作为后续甘氨酸的干预浓度。干预6.5 h,单纯烧伤血清组细胞AMP/ATP比值较正常血清组、1.2 mmol/L甘氨酸组、1.6 mmol/L甘氨酸组明显升高( P值均<0.05),1.6 mmol/L甘氨酸组细胞AMP/ATP比值较0.8 mmol/L甘氨酸组明显降低( P<0.05)。干预6.5 h,正常血清组、单纯烧伤血清组、0.8 mmol/L 甘氨酸组、1.2 mmol/L 甘氨酸组、1.6 mmol/L甘氨酸组细胞p-mTORC1、p-p70 S6K、p-4E-BP1蛋白表达水平分别为1.001±0.037、0.368±0.020、1.153±0.019、1.128±0.062、1.028±0.037,0.96±0.07、0.63±0.12、1.17±0.13、1.13±0.16、1.11±0.11,0.98±0.06、0.45±0.08、1.13±0.05、0.77±0.12、0.51±0.13。与单纯烧伤血清组比较,正常血清组与各甘氨酸组细胞p-mTORC1、p-p70 S6K和p-4E-BP1蛋白表达均明显升高( P<0.05),p-AMPK蛋白表达均明显降低( P<0.05);与0.8 mmol/L甘氨酸组比较,1.2 mmol/L甘氨酸组细胞p-4E-BP1蛋白表达以及1.6 mmol/L甘氨酸组细胞p-mTORC1与p-4E-BP1蛋白表达均明显降低( P<0.05);与1.2 mmol/L甘氨酸组比较,1.6 mmol/L甘氨酸组细胞p-mTORC1、p-4E-BP1蛋白表达均明显降低( P<0.05),p-AMPK蛋白表达明显升高( P<0.05)。与正常血清组比较,单纯烧伤血清组细胞干预1.5、3.5、6.5 h微管蛋白表达均明显降低( P<0.05),干预1.5、3.5 h HSP70表达和干预3.5、6.5 h MT表达均明显升高( P<0.05);单纯烧伤血清组与0.8 mmol/L甘氨酸+25 ng/mL雷帕霉素组细胞干预1.5、3.5、6.5 h HSP70、MT表达以及干预1.5、3.5 h微管蛋白表达均明显低于0.8 mmol/L甘氨酸组( P<0.05)。干预6.5 h,与正常血清组比较,单纯烧伤血清组细胞微管结构紊乱;0.8 mmol/L甘氨酸组细胞边界较单纯烧伤血清组清晰,微管结构近核部位排列整齐;与0.8 mmol/L甘氨酸组比较,0.8 mmol/L甘氨酸+25 ng/mL雷帕霉素组细胞边界不清,微管结构紊乱。 结论 烧伤血清可导致大鼠心肌细胞受损,甘氨酸可通过AMP活化蛋白激酶显著上调哺乳动物雷帕霉素靶蛋白/p70核糖体蛋白S6激酶/真核翻译起始因子4E结合蛋白1信号通路,促进心肌细胞保护性蛋白HSP70、MT和微管蛋白合成,稳定微管结构,实现心肌细胞保护作用。 Abstract:Objective To investigate the effect and mechanism of glycine on rat cardiomyocytes pretreated with serum from burned rats (hereinafter referred to as burn serum). Methods Experimental research methods were adopted. Thirty gender equally balanced Wistar rats aged 7 to 8 weeks were collected, 10 of which were used to prepare normal rat serum (hereinafter referred to as normal serum), and the other 20 were inflicted with full-thickness burn of 30% total body surface area to prepare burn serum. Primary cardiomyocytes were isolated and cultured from the apical tissue of 180 Wistar rats aged 1 to 3 days by either gender for follow-up experiments. Cells were divided into normal serum group and burn serum group treated with corresponding serum according to the random number table (the same grouping method below). Trypanosoma blue staining was performed at post treatment hour (PTH) 1, 3, 6, 9, and 12 to detect the cell survival rate. Cells were divided into burn serum alone group treated with burn serum for 6 h followed by routine culture of 30 min and 0.4 mmol/L glycine group, 0.8 mmol/L glycine group, 1.2 mmol/L glycine group, 1.6 mmol/L glycine group, and 2.0 mmol/L glycine group treated with burn serum for 6 h followed by culture of 30 min with corresponding final molarity of glycine, i.e., at post intervention hour (PIH) 6.5, the cell survival rate was detected as before. Cells were divided into normal serum group, burn serum alone group, 0.8 mmol/L glycine group, 1.2 mmol/L glycine group, and 1.6 mmol/L glycine group, with the same intervention of 6.5 h as before, respectively. The content of adenosine monophosphate (AMP) and adenosine triphosphate (ATP) was detected by high performance liquid chromatography, and the AMP/ATP ratio was calculated. The protein expressions of phosphorylated mammalian target of rapamycin complex 1 (p-mTORC1), phosphorylated p70 ribosomal protein S6 kinase (p-p70 S6K), phosphorylated eukaryotic translation initiation factor 4E-binding protein 1 (p-4E-BP1), and phosphorylated AMP-activated protein kinase (p-AMPK) were detected by Western blotting. Cells were divided into normal serum group, burn serum alone group, 0.8 mmol/L glycine group intervened as before and 0.8 mmol/L glycine+25 ng/mL rapamycin group treated with burn serum followed by culture with two reagents. The expressions of heat shock protein 70 (HSP70), metallothionein (MT), and tubulin were detected by immunofluorescence method after 30 min of corresponding culture at PTH 1, 3, and 6, i.e., at PIH 1.5, 3.5, and 6.5, and the microtubule morphology was observed at PIH 6.5. The sample number at each time point was 10. Data were statistically analyzed with analysis of variance for factorial design, one-way analysis of variance, least significant difference (LSD)- t test, LSD test, and Bonferroni correction. Results At PTH 1, 3, 6, 9, and 12, the cell survival rates in burn serum group were significantly lower than those in normal serum group (with t values of 4.96, 16.83, 35.51, 34.33, and 27.88, P<0.05). In burn serum group, the cell survival rate at PTH 3, 6, 9, or 12 was significantly lower than that at PTH 1 ( P<0.05), the cell survival rate at PTH 6, 9, or 12 was significantly lower than that at PTH 3 ( P<0.05), and the cell survival rate at PTH 6 was similar to that at PTH 9 ( P>0.05) but significantly higher than that at PTH 12 ( P<0.05). Treatment of 6 h was selected as the follow-up intervention time of burn serum. At PIH 6.5, compared with that in burn serum alone group, the cell survival rate in each glycine group was significantly increased ( P<0.05). The cell survival rate in 0.8 mmol/L glycine group was the highest, and 0.8, 1.2, and 1.6 mmol/L were selected as subsequent glycine intervention concentrations. At PIH 6.5, the AMP/ATP ratio of cells in burn serum alone group was significantly higher than that in normal serum group, 1.2 mmol/L glycine group, or 1.6 mmol/L glycine group ( P values all <0.05), and the AMP/ATP ratio of cells in 1.6 mmol/L glycine group was significantly lower than that in 0.8 mmol/L glycine group ( P<0.05). At PIH 6.5, the protein expressions of p-mTORC1, p-p70 S6K, and p-4E-BP1 of cells in normal serum group, burn serum alone group, 0.8 mmol/L glycine group, 1.2 mmol/L glycine group, and 1.6 mmol/L glycine group were 1.001±0.037, 0.368±0.020, 1.153±0.019, 1.128±0.062, 1.028±0.037, 0.96±0.07, 0.63±0.12, 1.17±0.13, 1.13±0.16, 1.11±0.11, and 0.98±0.06, 0.45±0.08, 1.13±0.05, 0.77±0.12, 0.51±0.13. Compared with those in burn serum alone group, the protein expressions of p-mTORC1, p-p70 S6K, and p-4E-BP1 of cells in normal serum group and each glycine group were significantly increased ( P<0.05), while the protein expressions of p-AMPK were significantly decreased ( P<0.05). Compared with those in 0.8 mmol/L glycine group, the protein expression of p-4E-BP1 of cells in 1.2 mmol/L glycine group and the protein expressions of p-mTORC1 and p-4E-BP1 of cells in 1.6 mmol/L glycine group were significantly decreased ( P<0.05). Compared with those in 1.2 mmol/L glycine group, the protein expressions of p-mTORC1 and p-4E-BP1 of cells in 1.6 mmol/L glycine group were significantly decreased ( P<0.05), while the protein expression of p-AMPK was significantly increased ( P<0.05). Compared with those in normal serum group, the expression of tubulin of cells in burn serum alone group was significantly decreased at PIH 1.5, 3.5, and 6.5 ( P<0.05), while the expression of HSP70 of cells at PIH 1.5 and 3.5 and the expression of MT at PIH 3.5 and 6.5 were significantly increased ( P<0.05). The expressions of HSP70 and MT of cells at PIH 1.5, 3.5, and 6.5 and the expression of tubulin at PIH 1.5 and 3.5 in burn serum alone group and 0.8 mmol/L glycine+25 ng/mL rapamycin group were significantly lower than those in 0.8 mmol/L glycine group ( P<0.05). At PIH 6.5, compared with that in normal serum group, the cell microtubule structure in burn serum alone group was disordered; the cell boundary in 0.8 mmol/L glycine group was clearer than that in burn serum alone group, and the microtubule structure arranged neatly near the nucleus. Compared with that in 0.8 mmol/L glycine group, 0.8 mmol/L glycine+25 ng/mL rapamycin group had unclear cell boundaries and disordered microtubule structure. Conclusions Burn serum can cause cardiomyocytes damage in rats. Glycine can significantly up-regulate mammalian target of rapamycin/p70 ribosomal protein S6 kinase/eukaryotic translation initiation factor 4E-binding protein 1 signaling pathway through AMP-activated protein kinase, promote the synthesis of protective proteins HSP70, MT, and tubulin, stabilize the microtubule structure, and exert cardiomyocytes protection function. -
参考文献
(28) [1] 黄跃生.严重烧伤早期心肌损害机制及临床意义的再认识[J].中华烧伤杂志,2016,32(5):257-259.DOI: 10.3760/cma.j.issn.1009-2587.2016.05.001. [2] ClarkA,ImranJ,MadniT,et al.Nutrition and metabolism in burn patients[J/OL].Burns Trauma,2017,5:11[2023-02-06].https://pubmed.ncbi.nlm.nih.gov/28428966/.DOI: 10.1186/s41038-017-0076-x. [3] 刘琰,王际壮.烧伤应激反应及其调控策略[J].中华烧伤杂志,2021,37(2):126-130.DOI: 10.3760/cma.j.cn501120-20201125-00499. [4] 彭曦.烧伤临床营养新视角[J].中华烧伤杂志,2019,35(5):321-325.DOI: 10.3760/cma.j.issn.1009-2587.2019.05.001. [5] 吕尚军,范荣辉,吴丹,等.谷氨酰胺对经烧伤大鼠血清干预的大鼠心肌细胞的作用及其细胞信号机制[J].中华烧伤杂志,2021,37(12):1149-1157.DOI: 10.3760/cma.j.cn501120-20210601-00208. [6] ZhangY,LvSJ,YanH,et al.Effects of glycine supplementation on myocardial damage and cardiac function after severe burn[J].Burns,2013,39(4):729-735.DOI: 10.1016/j.burns.2012.09.006. [7] MorioA,TsutsumiR,KondoT,et al.Leucine induces cardioprotection in vitro by promoting mitochondrial function via mTOR and Opa-1 signaling[J].Nutr Metab Cardiovasc Dis,2021,31(10):2979-2986.DOI: 10.1016/j.numecd.2021.06.025. [8] Ley-NgardigalS,BertolinG.Approaches to monitor ATP levels in living cells: where do we stand?[J].FEBS J,2022,289(24):7940-7969.DOI: 10.1111/febs.16169. [9] Costa-MattioliM,WalterP.The integrated stress response: from mechanism to disease[J].Science,2020,368(6489):eaat5314.DOI: 10.1126/science.aat5314. [10] OsborneT,WallB,EdgarDW,et al.Current understanding of the chronic stress response to burn injury from human studies[J/OL].Burns Trauma,2023,11:tkad007[2023-02-06].https://pubmed.ncbi.nlm.nih.gov/36926636/.DOI: 10.1093/burnst/tkad007. [11] SomeroGN.The cellular stress response and temperature: function, regulation, and evolution[J].J Exp Zool A Ecol Integr Physiol,2020,333(6):379-397.DOI: 10.1002/jez.2344. [12] LiuP,WangMZ,ZhongWX,et al.Stress-responsive genes (hsp70 and mt) and genotoxicity elicited by roxarsone exposure in Carassius auratus[J].Environ Toxicol Pharmacol,2018,62:132-139.DOI: 10.1016/j.etap.2018.07.004. [13] ZhongZ,WheelerMD,LiX,et al.L-glycine: a novel antiinflammatory, immunomodulatory, and cytoprotective agent[J].Curr Opin Clin Nutr Metab Care,2003,6(2):229-240.DOI: 10.1097/00075197-200303000-00013. [14] PanS,FanM,LiuZ,et al.Serine, glycine and one-carbon metabolism in cancer (Review)[J].Int J Oncol,2021,58(2):158-170.DOI: 10.3892/ijo.2020.5158. [15] GutP,LizzoG,MigliavaccaE,et al. Effects of glycine and n-acetylcysteine on glutathione levels and mitochondrial energy metabolism in healthy aging[J].Innov Aging,2021,5(Suppl 1):S685.DOI: 10.1093/geroni/igab046.2574. [16] SaxtonRA,SabatiniDM.mTOR signaling in growth, metabolism, and disease[J].Cell,2017,168(6):960-976.DOI: 10.1016/j.cell.2017.02.004. [17] ParkJH,LeeG,BlenisJ.Structural insights into the activation of mTORC1 on the lysosomal surface[J].Trends Biochem Sci,2020,45(5):367-369.DOI: 10.1016/j.tibs.2020.02.004. [18] Francois-VaughanH,AdebayoAO,BrilliantKE,et al.Persistent effect of mTOR inhibition on preneoplastic foci progression and gene expression in a rat model of hepatocellular carcinoma[J].Carcinogenesis,2016,37(4):408-419.DOI: 10.1093/carcin/bgw016. [19] LinSC,HardieDG.AMPK: sensing glucose as well as cellular energy status[J].Cell Metab,2018,27(2):299-313.DOI: 10.1016/j.cmet.2017.10.009. [20] SteinbergGR,CarlingD.AMP-activated protein kinase: the current landscape for drug development[J].Nat Rev Drug Discov,2019,18(7):527-551.DOI: 10.1038/s41573-019-0019-2. [21] ZhuJ,WangYF,ChaiXM,et al.Exogenous NADPH ameliorates myocardial ischemia-reperfusion injury in rats through activating AMPK/mTOR pathway[J].Acta Pharmacol Sin,2020,41(4):535-545.DOI: 10.1038/s41401-019-0301-1. [22] XiangF,MaSY,LvYL,et al.Tumor necrosis factor receptor-associated protein 1 regulates hypoxia-induced apoptosis through a mitochondria-dependent pathway mediated by cytochrome c oxidase subunit II[J/OL].Burns Trauma,2019,7:16[2023-02-06].https://pubmed.ncbi.nlm.nih.gov/31143823/.DOI: 10.1186/s41038-019-0154-3. [23] WangW,WuZ,DaiZ,et al.Glycine metabolism in animals and humans: implications for nutrition and health[J].Amino Acids,2013,45(3):463-477.DOI: 10.1007/s00726-013-1493-1. [24] BuruteM,KapiteinLC.Cellular logistics: unraveling the interplay between microtubule organization and intracellular transport[J].Annu Rev Cell Dev Biol,2019,35:29-54.DOI: 10.1146/annurev-cellbio-100818-125149. [25] YuanAT,KorkolaNC,WongDL,et al.Metallothionein Cd4S11 cluster formation dominates in the protection of carbonic anhydrase[J].Metallomics,2020,12(5):767-783.DOI: 10.1039/d0mt00023j. [26] 谢果,陈伟,王海英.金属硫蛋白在心肌缺血/再灌注损伤中保护作用的研究进展[J].国际麻醉学与复苏杂志,2021,42(9):1004-1008.DOI: 10.3760/cma.j.cn321761-20200601-00382. [27] YangL,MaJ,TanY,et al.Cardiac-specific overexpression of metallothionein attenuates L-NAME-induced myocardial contractile anomalies and apoptosis[J].J Cell Mol Med,2019,23(7):4640-4652.DOI: 10.1111/jcmm.14375. [28] KuznetsovAV,JavadovS,GrimmM,et al.Crosstalk between mitochondria and cytoskeleton in cardiac cells[J].Cells,2020,9(1):222.DOI: 10.3390/cells9010222. -

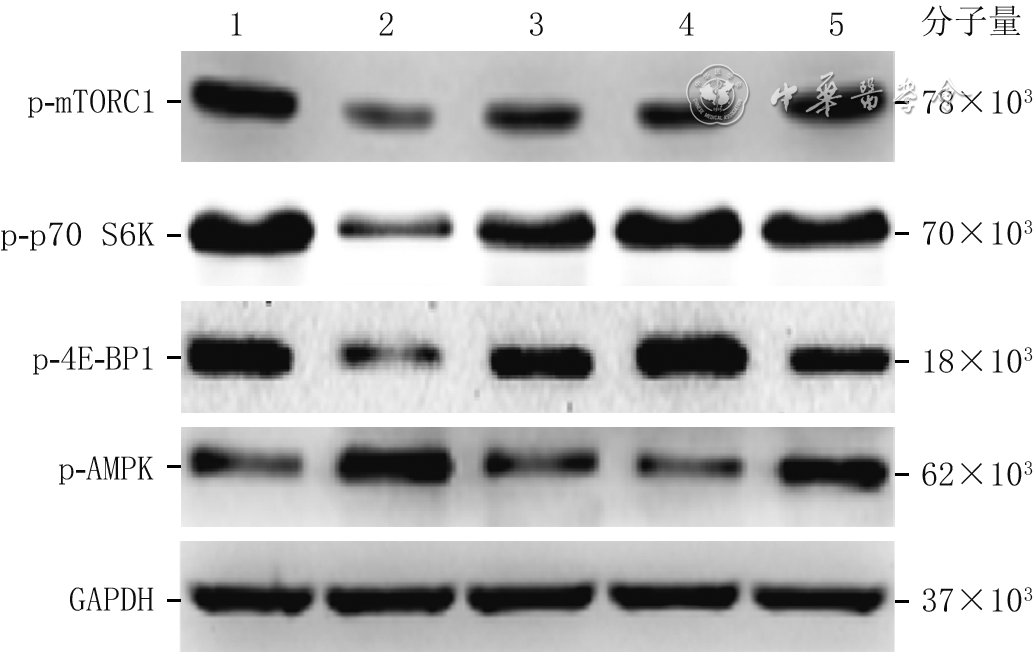
1 蛋白质印迹法检测的5组大鼠心肌细胞干预6.5 h时哺乳动物雷帕霉素靶蛋白信号通路上下游分子蛋白表达
注:正常血清组与单纯烧伤血清组细胞分别用正常大鼠血清与烧伤大鼠血清处理6 h并常规培养30 min,3个甘氨酸组细胞均用烧伤大鼠血清处理6 h并加相应终物质的量浓度甘氨酸培养30 min;p-mTORC1为磷酸化哺乳动物雷帕霉素靶蛋白复合物1,p-p70 S6K为磷酸化p70核糖体蛋白S6激酶,p-4E-BP1为磷酸化真核翻译起始因子4E结合蛋白1,p-AMPK为磷酸化腺苷一磷酸活化蛋白激酶,GAPDH为3-磷酸甘油醛脱氢酶;条带上方1、2、3、4、5分别指示正常血清组、单纯烧伤血清组、0.8 mmol/L甘氨酸组、1.2 mmol/L甘氨酸组、1.6 mmol/L甘氨酸组

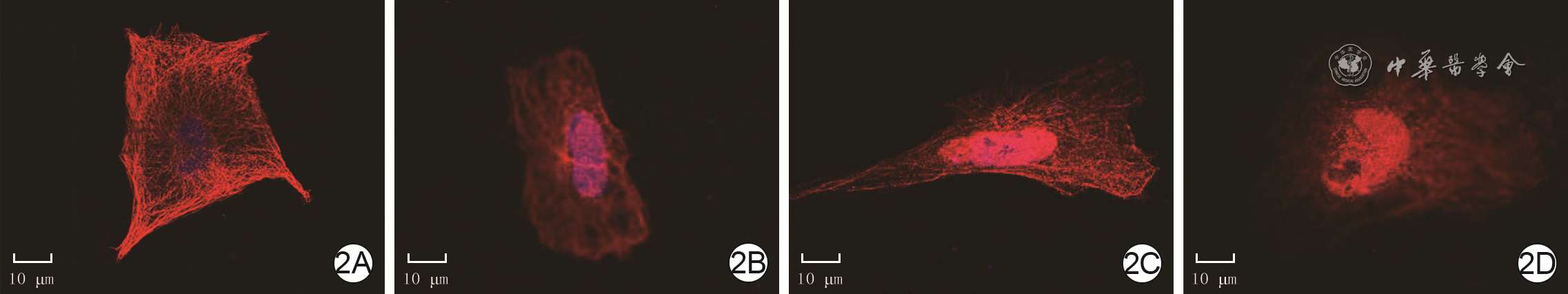
2 4组大鼠心肌细胞干预6.5 h微管形态 异硫氰酸荧光素-Hoechst 33342×800。2A.正常血清组细胞呈多边形,微管结构完整;2B.单纯烧伤血清组细胞皱缩呈圆形,微管结构紊乱;2C.0.8 mmol/L甘氨酸组微管破坏程度较图2B有所减轻;2D.0.8 mmol/L甘氨酸+25 ng/mL雷帕霉素组微管破坏程度比图2C严重
注:正常血清组与单纯烧伤血清组细胞分别用正常大鼠血清与烧伤大鼠血清处理6 h并常规培养30 min,另2组细胞均用烧伤大鼠血清处理6 h并加甘氨酸或甘氨酸+雷帕霉素培养30 min;心肌细胞微管阳性染色为红色,细胞核阳性染色为蓝色
表1 2组大鼠心肌细胞处理各时间点存活率比较(%,
组别 样本数 1 h 3 h 6 h 9 h 12 h F值 P值 正常血清组 10 90.0±3.0 88.3±3.4 91.8±2.7 90.9±3.1 90.4±1.3 1.60 0.089 烧伤血清组 10 83.3±5.6 61.3±4.4 a 44.9±2.9 ab 42.6±2.8 ab 41.2±3.7 abc 241.32 <0.001 t值 4.96 16.83 35.51 34.33 27.88 P值 <0.001 <0.001 <0.001 <0.001 <0.001 注:血清均来自大鼠;处理因素主效应, F=1 931.77, P<0.001;时间因素主效应, F=141.62, P<0.001;两者交互作用, F=135.76, P<0.001;与组内1 h比较, a P<0.05;与组内3 h比较, b P<0.05;与组内6 h比较, c P<0.05  下载: 导出CSV
下载: 导出CSV
表2 5组大鼠心肌细胞干预6.5 h时哺乳动物雷帕霉素靶蛋白信号通路上下游分子蛋白表达比较(
组别 样本数 p-mTORC1 p-p70 S6K p-4E-BP1 p-AMPK 正常血清组 10 1.001±0.037 0.96±0.07 0.98±0.06 0.42±0.08 单纯烧伤血清组 10 0.368±0.020 a 0.63±0.12 a 0.45±0.08 a 0.88±0.08 a 0.8 mmol/L甘氨酸组 10 1.153±0.019 b 1.17±0.13 b 1.13±0.05 b 0.73±0.09 b 1.2 mmol/L甘氨酸组 10 1.128±0.062 b 1.13±0.16 b 0.77±0.12 bc 0.63±0.11 b 1.6 mmol/L甘氨酸组 10 1.028±0.037 bcd 1.11±0.11 b 0.51±0.13 bcd 0.76±0.07 bd F值 706.31 34.47 84.14 39.88 P值 <0.001 <0.001 <0.001 <0.001 注:正常血清组与单纯烧伤血清组细胞分别用正常大鼠血清与烧伤大鼠血清处理6 h并常规培养30 min,3个甘氨酸组细胞均用烧伤大鼠血清处理6 h并加相应终物质的量浓度甘氨酸培养30 min;p-mTORC1为磷酸化哺乳动物雷帕霉素靶蛋白复合物1,p-p70 S6K为磷酸化p70核糖体蛋白S6激酶,p-4E-BP1为磷酸化真核翻译起始因子4E结合蛋白1,p-AMPK为磷酸化腺苷一磷酸活化蛋白激酶;与正常血清组比较, a P<0.05;与单纯烧伤血清组比较, b P<0.05;与0.8 mmol/L甘氨酸组比较, c P<0.05;与1.2 mmol/L甘氨酸组比较, d P<0.05
下载: 导出CSV
表3 4组大鼠心肌细胞干预各时间点HSP70与MT和微管蛋白表达比较(
组别与时间点 样本数 HSP70 MT 微管蛋白 正常血清组 30 1.5 h 3.63±1.26 5.5±0.9 16.2±1.4 3.5 h 3.52±0.35 5.6±1.0 15.7±1.2 6.5 h 3.44±0.39 5.5±0.7 15.5±1.1 单纯烧伤血清组 30 1.5 h 7.51±0.91 a 5.2±1.4 9.0±1.5 a 3.5 h 5.44±1.13 a 7.4±1.0 a 6.8±1.8 a 6.5 h 4.11±0.79 8.4±0.6 a 6.6±1.3 a 0.8 mmol/L甘氨酸组 30 1.5 h 9.73±1.19 b 11.3±0.7 b 10.9±1.3 b 3.5 h 12.37±1.43 b 12.7±1.8 b 7.9±1.3 b 6.5 h 15.21±1.14 b 16.2±1.3 b 7.2±1.1 0.8 mmol/L甘氨酸+25 ng/mL雷帕霉素组 30 1.5 h 1.34±0.29 c 3.1±1.1 c 7.8±1.4 c 3.5 h 1.69±0.49 c 2.9±0.4 c 6.3±0.7 c 6.5 h 2.28±0.73 c 2.8±1.2 c 6.4±1.7 F 1值 149.30 103.30 71.10 P 1值 <0.001 <0.001 <0.001 F 2值 191.22 125.93 108.17 P 2值 <0.001 <0.001 <0.001 F 3值 285.03 337.95 107.69 P 3值 <0.001 <0.001 <0.001 注:正常血清组与单纯烧伤血清组细胞分别用正常大鼠血清与烧伤大鼠血清处理6 h并常规培养30 min,后2组细胞均用烧伤大鼠血清处理6 h并加甘氨酸或甘氨酸+雷帕霉素培养30 min;HSP70为热休克蛋白70,MT为金属硫蛋白;各组各时间点样本数均为10;HSP70、MT和微管蛋白处理因素主效应, F值分别为493.27、479.79、720.29, P值均<0.001;时间因素主效应, F值分别为8.68、31.59、24.34, P值均<0.001;两者交互作用, F值分别为30.29、13.84、2.42, P值均<0.001; F 1值、 P 1值, F 2值、 P 2值, F 3值、 P 3值分别为4组细胞干预1.5、3.5、6.5 h各指标总体比较所得;与正常血清组比较, a P<0.05;与单纯烧伤血清组比较, b P<0.05;与0.8 mmol/L甘氨酸组比较, c P<0.05
下载: 导出CSV
-

 下载:
下载:


计量
- 文章访问数: 2271
- HTML全文浏览量: 175
- PDF下载量: 11
- 被引次数: 0
