



Exploring the causality between intestinal flora and hyperplastic scars of human based on two-sample Mendelian randomization analysis
-
摘要:
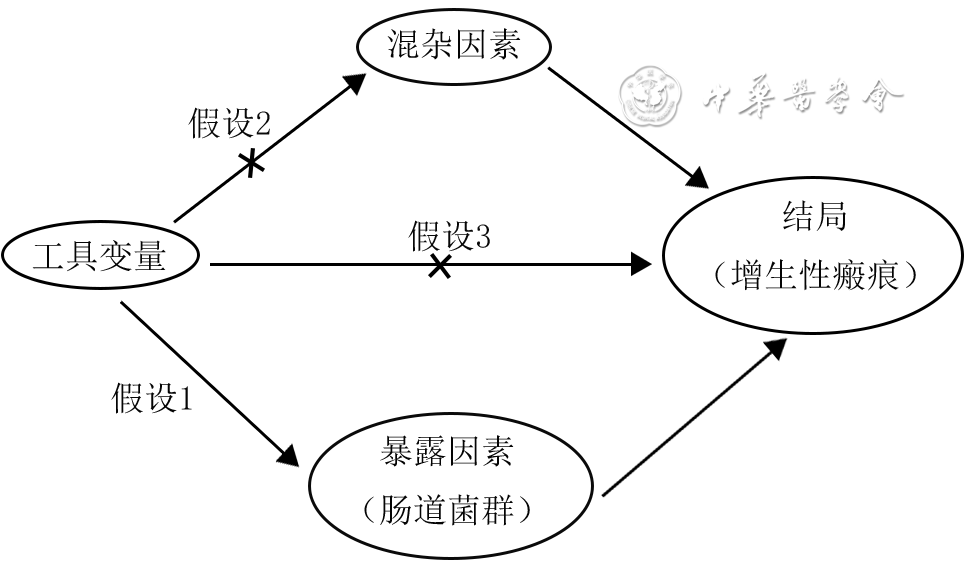
目的 探究人肠道菌群与增生性瘢痕(HS)之间的因果关系。 方法 该研究为基于双样本孟德尔随机化(TSMR)分析的研究。从全基因组关联分析数据库获得人肠道菌群(18 473个样本)和HS(208 248个样本)的数据。提取已知肠道菌群门、纲、目、科和属5个水平的遗传变异基因,即单核苷酸多态性(SNP)作为工具变量,并进行连锁不平衡(LD)分析。使用PhenoScanner V2数据库进行人类基因型-表型关联分析,排除与HS不相关的肠道菌群SNP并分析获得的SNP是否为弱工具变量。采用TSMR分析的4种方法,即逆方差加权(IVW)、MR-Egger回归、加权中位数和加权模式,对肠道菌群SNP与HS之间的因果关系进行分析。绘制前述4种分析方法得出重要结果的散布图,分析肠道菌群SNP与HS的相关性。采用IVW检验和MR-Egger回归检验评估肠道菌群SNP的异质性,采用MR-Egger回归检验和MR-PRESSO离群值检验评估肠道菌群SNP的水平多效性,采用留一敏感性分析确定HS是否由肠道菌群中的单个SNP引起。对HS SNP与肠杆菌属或HS SNP与瘤胃球菌2属分别进行逆向TSMR分析,检测它们之间是否存在反向因果关系。 结果 共获得196个已知肠道菌群,属于9门、16纲、20目、32科、119属,从每个菌群中均获得多个SNP作为工具变量。LD分析显示,除rs1000888、rs12566247、rs994794外,其余肠道菌群SNP均符合遗传变异与暴露因素密切相关的假设。人类基因型-表型关联分析显示,经LD分析后获得的SNP均未被排除且均不是弱工具变量。IVW、MR-Egger回归、加权中位数和加权模式的TSMR分析显示,肠杆菌属及瘤胃球菌2属均与HS存在因果关系。其中,IVW和MR-Egger回归的森林图分析还显示,肠杆菌属的16个SNP(该菌属SNP个数下同)和瘤胃球菌2属的15个SNP(该菌属SNP个数下同)均为HS的保护因素。进一步地,IVW分析显示肠杆菌属SNP(比值比为0.62,95%置信区间为0.41~0.93, P<0.05)和瘤胃球菌2属SNP(比值比为0.62,95%置信区间为0.40~0.97, P<0.05)均与HS发生风险呈负相关。散布图显示,肠杆菌属和瘤胃球菌2属的SNP均是HS的保护因素。IVW检验和MR-Egger回归检验均显示,肠杆菌属SNP( Q值分别为5.73、5.76, P>0.05)和瘤胃球菌2属SNP( Q值分别为13.67、15.61, P>0.05)均无异质性。MR-Egger回归检验显示,肠杆菌属和瘤胃球菌2属的SNP均无水平多效性(截距分别为0.01、0.06, P>0.05);MR-PRESSO离群值检验显示,肠杆菌属和瘤胃球菌2属的SNP均无水平多效性( P>0.05)。留一敏感性分析显示,无单个肠道菌群SNP驱动HS的发生。逆向TSMR分析显示,HS SNP与肠杆菌属之间、HS SNP与瘤胃球菌2属之间均不存在任何反向因果关系(比值比分别为1.01、0.99,95%置信区间分别为0.97~1.06、0.96~1.04, P>0.05)。 结论 人肠道菌群与HS之间存在一定因果关系,肠杆菌属及瘤胃球菌2属对抑制HS有一定作用。 Abstract:Objective To investigate the causality between intestinal flora and hypertrophic scars (HS) of human. Methods This study was a study based on two-sample Mendelian randomization (TSMR) analysis. The data on intestinal flora ( n=18 473) and HS ( n=208 248) of human were obtained from the genome-wide association study database. Genetically variable genes at five levels (phylum, class, order, family, and genus) of known intestinal flora, i.e., single nucleotide polymorphisms (SNPs), were extracted as instrumental variables for linkage disequilibrium (LD) analysis. Human genotype-phenotype association analysis was performed using PhenoScanner V2 database to exclude SNPs unrelated to HS in intestinal flora and analyze whether the selected SNPs were weak instrumental variables. The causal relationship between intestinal flora SNPs and HS was analyzed through four methods of TSMR analysis, namely inverse variance weighted (IVW), MR-Egger regression, weighted median, and weighted mode. Scatter plots of significant results from the four aforementioned analysis methods were plotted to analyze the correlation between intestinal flora SNPs and HS. Both IVW test and MR-Egger regression test were used to assess the heterogeneity of intestinal flora SNPs, MR-Egger regression test and MR-PRESSO outlier test were used to assess the horizontal multiplicity of intestinal flora SNPs, and leave-one-out sensitivity analysis was used to determine whether HS was caused by a single SNP in the intestinal flora. Reverse TSMR analyses were performed for HS SNPs and genus Intestinimonas or genus Ruminococcus2, respectively, to detect whether there was reverse causality between them. Results A total of 196 known intestinal flora, belonging to 9 phyla, 16 classes, 20 orders, 32 families, and 119 genera, were obtained, and multiple SNPs were obtained from each flora as instrumental variables. LD analysis showed that the SNPs of the intestinal flora were consistent with the hypothesis that genetic variation was strongly associated with exposure factors, except for rs1000888, rs12566247, and rs994794. Human genotype-phenotype association analysis showed that none of the selected SNPs after LD analysis was excluded and there were no weak instrumental variables. IVW, MR-Egger regression, weighted median, and weighted mode of TSMR analysis showed that both genus Intestinimonas and genus Ruminococcus2 were causally associated with HS. Among them, forest plots of IVW and MR-Egger regression analyses also showed that 16 SNPs (the same SNPs number of this genus below) of genus Intestinimonas and 15 SNPs (the same SNPs number of this genus below) of genus Ruminococcus2 were protective factors for HS. Further, IVW analysis showed that genus Intestinimonas SNPs (with odds ratio of 0.62, 95% confidence interval of 0.41-0.93, P<0.05) and genus Ruminococcus2 SNPs (with odds ratio of 0.62, 95% confidence interval of 0.40-0.97, P<0.05) were negatively correlated with the risk of HS. Scatter plots showed that SNPs of genus Intestinimonas and genus Ruminococcus2 were protective factors of HS. Both IVW test and MR-Egger regression test showed that SNPs of genus Intestinimonas (with Q values of 5.73 and 5.76, respectively, P>0.05) and genus Ruminococcus2 (with Q values of 13.67 and 15.61, respectively, P>0.05) were not heterogeneous. MR-Egger regression test showed that the SNPs of genus Intestinimonas and genus Ruminococcus2 had no horizontal multiplicity (with intercepts of 0.01 and 0.06, respectively, P>0.05); MR-PRESSO outlier test showed that the SNPs of genus Intestinimonas and genus Ruminococcus2 had no horizontal multiplicity ( P>0.05). Leave-one-out sensitivity analysis showed that no single intestinal flora SNP drove the occurrence of HS. Reverse TSMR analysis showed no reverse causality between HS SNPs and genus Intestinimonas or genus Ruminococcus2 (with odds ratios of 1.01 and 0.99, respectively, 95% confidence intervals of 0.97-1.06 and 0.96-1.04, respectively, P>0.05). Conclusions There is a causal relationship between intestinal flora and HS of human, in which genus Intestinimonas and genus Ruminococcus2 have a certain effect on inhibiting HS. -
创面是临床常见病症[1],创面血管新生是创面修复的关键环节之一[2]。血管新生是一个受到精确调控的病理生理过程[3],该过程涉及多种细胞与细胞因子[4, 5],其中微血管内皮细胞是参与此过程的主要功能细胞[6],它们在多种细胞因子及信号通路的精确调控下,通过迁移、增殖、芽生等形成新生血管[7, 8, 9]。
P311,官方基因符号NERP,定位于人5号染色体和小鼠18号染色体上,编码分子量为8×103的胞内蛋白,该蛋白是一种在种属间高度保守的多功能蛋白[10]。研究表明,P311蛋白在组织纤维化[11, 12, 13, 14]、神经系统疾病[15]、肿瘤浸润[16, 17]、血压维持[18]、组织再生[19, 20]等多方面发挥重要作用。近年来研究表明,P311在创面愈合中发挥重要作用[21, 22, 23]。本课题组前期研究表明,人皮肤微血管内皮细胞在静息状态下低表达P311,在炎症刺激条件下高表达P311,进而参与创面血管新生[22,24],但目前P311参与血管新生的机制仍不清楚。因此,本课题组通过前期构建的P311腺病毒转染人微血管内皮细胞1(HMEC-1),使之高表达P311,从而模拟创面愈合过程中P311的表达模式,进而探讨P311对HMEC-1体外血管形成能力的影响及其可能的分子机制。
1. 材料与方法
1.1 主要试剂和仪器来源
基质胶购自美国Corning公司,HMEC系购自美国典型培养物保藏中心,MCDB131培养基购自美国Gibco公司,二甲基亚砜(DMSO)购自德国Meck公司,脂质体2000购自北京索莱宝科技有限公司,细胞计数试剂盒8(CCK-8)、二辛丁酸蛋白浓度测定试剂盒购自海门市碧云天生物技术研究所,VEGF受体2(VEGFR2)、小干扰RNA(siRNA)和阴性对照siRNA均购自广州锐博生物科技有限公司,胞外信号调节激酶1/2(ERK1/2)抑制剂SCH772984购自上海蓝木化工有限公司,兔抗小鼠VEGFR2单克隆抗体、兔抗小鼠磷酸化VEGFR2单克隆抗体、兔抗小鼠ERK1/2单克隆抗体、兔抗小鼠磷酸化ERK1/2单克隆抗体购自美国Cell Signaling Technology公司,小鼠源性GAPDH单克隆抗体购自上海康成生物工程有限公司,辣根过氧化物酶标记的山羊抗兔IgG多克隆抗体和山羊抗小鼠IgG多克隆抗体均购自北京中杉金桥生物技术有限公司。710 型多功能酶标仪购自美国Thermo Fisher公司,LSM510 Meta型活细胞工作站购自德国Zeiss公司,IX71型倒置相差显微镜购自日本Olympus公司,ChemiDoc XRS型凝胶成像仪购自美国Bio-Rad公司。
1.2 HMEC-1的培养和传代
将含有10 ng/mL EGF、10 mmol/L谷氨酰胺、1 µg/mL氢化可的松和体积分数10%胎牛血清的MCDB131培养基(以下称完全培养基)加入底面积25 cm2的培养瓶后,将1×106个HMEC-1接种至该培养瓶中培养,每隔2 d换液1次,待细胞生长达85%融合以上,使用2.5 g/L胰蛋白酶+0.1 g/L乙二胺四乙酸消化后,按1∶3进行细胞传代。取第3~5代细胞用于后续实验。
1.3 P311高表达对HMEC-1增殖、迁移和血管形成能力的影响
1.3.1 细胞分组及处理
用完全培养基调整HMEC-1浓度为1×105个/mL,接种于6孔板中,每孔1 mL。待细胞生长达60%左右融合时,将其按随机数字表法(分组方法下同)分为P311腺病毒组和空载腺病毒组。P311腺病毒组细胞每孔加入10 μL P311腺病毒,空载腺病毒组细胞每孔加入10 μL空载腺病毒(P311腺病毒和空载腺病毒均由本课题组前期构建),转染48 h后,收集细胞用于后续实验。
1.3.2 CCK-8法检测P311高表达对HMEC-1增殖活性的影响
取P311腺病毒组和空载腺病毒组细胞,用完全培养基调整细胞浓度为2×104个/mL,接种于96孔板中,每孔100 μL,每组3个复孔。于培养1、3、5 d,分别使用CCK-8法检测2组细胞增殖活性。检测时,吸弃原有培养基,将完全培养基和CCK-8溶液按体积比10∶1混合,每孔加入混合液100 μL,37 ℃孵育2 h,使用多功能酶标仪测定波长450 nm处的吸光度值,以此代表细胞增殖活性。本实验重复6次。
1.3.3 划痕试验检测P311高表达对HMEC-1迁移能力的影响
取P311腺病毒组和空载腺病毒组细胞,用完全培养基调整细胞浓度为2×104个/mL,接种于24孔板中,每孔1 mL,每组3个复孔。待细胞生长达90%以上融合时,吸弃原有培养基,加入500 μL含2.5 μg/mL丝裂霉素C的完全培养基继续培养2 h。使用100 μL移液枪枪头和直尺在HMEC-1单细胞层上做一直线划痕。吸弃原有培养基,加入PBS冲洗2遍,加入新鲜完全培养基。将细胞置于活细胞工作站倒置相差显微镜200倍镜下观察11 h,记录细胞迁移情况。使用ImageJ 1.48V图像分析软件(美国国立卫生研究院)测量划痕后0(即刻)、6、11 h剩余划痕面积,并计算剩余划痕面积百分比。剩余划痕面积百分比=剩余划痕面积÷初始划痕面积×100%。本实验重复6次。
1.3.4 体外血管形成实验检测P311高表达对HMEC-1血管形成能力的影响
首先将50 μL基质胶加入预冷的96孔板中,在37 ℃培养箱中放置45 min。然后加入用无血清MCDB131培养基培养24 h的P311腺病毒组和空载腺病毒组细胞,用完全培养基调整细胞浓度为4×105个/mL,每孔50 μL,每组3个复孔,在37 ℃培养箱中培养8 h,倒置相差显微镜200倍镜下观察血管形成情况并拍照,采用ImageJ 1.48V图像分析软件测量管状结构节点数和总长度。本实验重复6次。
1.3.5 蛋白质印迹法检测P311高表达对VEGFR2/ERK1/2信号通路的影响
取P311腺病毒组和空载腺病毒组细胞,充分裂解后, 以离心半径10 cm、14 000 r/min离心15 min后收集上清液,二辛丁酸法进行蛋白定量。向上样孔内加入20 μg待测蛋白质样品,十二烷基硫酸钠-聚丙烯酰胺凝胶电泳,湿法转膜后,用30 g/L牛血清白蛋白溶液封闭3 h,分别加入兔抗小鼠VEGFR2单克隆一抗、兔抗小鼠磷酸化VEGFR2单克隆一抗、兔抗小鼠ERK1/2单克隆一抗、兔抗小鼠磷酸化ERK1/2单克隆一抗(稀释比均为1∶1 000)及小鼠源性GAPDH单克隆一抗(稀释比为1∶10 000),4 ℃孵育过夜。次日取出抗体孵育盒,室温下静置1 h,洗涤后分别加入辣根过氧化物酶标记的山羊抗兔IgG多克隆二抗和山羊抗小鼠IgG多克隆二抗(稀释比均为1∶2 000),室温孵育1 h。化学发光液浸泡条带后,凝胶成像仪下曝光,并采集图像,Quantity one软件进行灰度值分析,VEGFR2、ERK1/2的蛋白表达量分别以其与GAPDH灰度值比值表示,磷酸化VEGFR2、磷酸化ERK1/2的蛋白表达量分别以其与VEGFR2、ERK1/2的灰度值比值表示。本实验重复6次。
1.4 P311高表达对HMEC-1 VEGFR2/ERK1/2信号通路及体外血管形成能力的影响
1.4.1 蛋白质印迹法检测siRNA-VEGFR2对P311高表达引起的VEGFR2/ERK1/2信号通路活化的影响
用完全培养基调整HMEC-1浓度为1×105个/mL,接种于6孔板中,每孔1 mL。待细胞生长达60%左右融合时,分为P311腺病毒+阴性对照siRNA组、空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组和空载腺病毒+siRNA-VEGFR2组,每组3个复孔。P311腺病毒+阴性对照siRNA组、空载腺病毒+阴性对照siRNA组细胞每孔分别加入10 μL P311腺病毒、空载腺病毒,24 h后,再采用脂质体2000转染终物质的量浓度50 nmol/L阴性对照siRNA;P311腺病毒+siRNA-VEGFR2组和空载腺病毒+siRNA-VEGFR2组细胞每孔分别加入10 μL P311腺病毒、空载腺病毒,24 h后,采用脂质体2000转染终物质的量浓度50 nmol/L的siRNA-VEGFR2(靶基因序列为5'-GGAAATCTCTTGCAAGCTA-3')。转染24 h,同1.3.5采用蛋白质印迹法检测VEGFR2、磷酸化VEGFR2、ERK1/2、磷酸化ERK1/2的蛋白表达情况。本实验重复6次。
1.4.2 体外血管形成实验检测siRNA-VEGFR2对P311高表达介导的血管形成作用的影响
取HMEC-1,同1.4.1分组及处理,同1.3.4检测各组细胞体外血管形成情况。本实验重复6次。
1.4.3 蛋白质印迹法检测ERK1/2抑制剂对P311高表达引起的ERK1/2信号通路活化的影响
取HMEC-1,用完全培养基调整HMEC-1浓度为1×105个/mL,接种于6孔板中,每孔1 mL,待细胞生长达60%左右融合时,分为P311腺病毒+DMSO组、载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组和空载腺病毒+ERK1/2抑制剂组,每组1孔。P311腺病毒+DMSO组和空载腺病毒+DMSO组细胞每孔分别加入10 μL P311腺病毒、空载腺病毒,24 h后,加入10 μL DMSO处理2 h;P311腺病毒+ERK1/2抑制剂组和空载腺病毒+ERK1/2抑制剂组细胞每孔分别加入10 μL P311腺病毒、空载腺病毒,24 h后,加入终物质的量浓度5 nmol/L ERK1/2抑制剂SCH772984处理2 h。 同1.3.5采用蛋白质印迹法检测ERK1/2和磷酸化ERK1/2的蛋白表达情况。本实验重复6次。
1.4.4 体外血管形成实验检测ERK1/2抑制剂对P311高表达介导的血管形成作用的影响
取HMEC-1,同1.4.3分组及处理,同1.3.4检测各组细胞体外血管形成情况。本实验重复6次。
1.5 统计学处理
采用SPSS 23.0统计软件进行数据分析。所有计量资料数据均符合正态分布,以
2. 结果
2.1 P311高表达对HMEC-1增殖、迁移和血管形成能力的影响
2.1.1 细胞增殖活性
与空载腺病毒组比较,P311腺病毒组细胞培养各时间点增殖活性均没有明显改变(P>0.05),见表1。
表1 2组人微血管内皮细胞1培养各时间点增殖活性比较(组别 样本数 1 d 3 d 5 d P311腺病毒组 6 0.188±0.030 0.415±0.032 0.713±0.030 空载腺病毒组 6 0.185±0.019 0.392±0.030 0.687±0.031 t值 -0.23 -1.30 -1.52 P值 0.822 0.222 0.160 注:处理因素主效应,F=5.10,P=0.074;时间因素主效应,F=720.12,P<0.001;两者交互作用,F=0.81,P=0.473 2.1.2 细胞迁移能力
P311腺病毒组细胞划痕后6、11 h剩余划痕面积百分比较空载腺病毒组明显降低(P<0.05),见表2、图1。
表2 2组人微血管内皮细胞1划痕后各时间点剩余划痕面积百分比比较(%,组别 样本数 6 h 11 h P311腺病毒组 6 47±7 9±5 空载腺病毒组 6 55±5 20±10 t值 -2.47 -2.62 P值 0.033 0.025 注:初始剩余划痕面积百分比均为100%;处理因素主效应,F=18.42,P=0.008;时间因素主效应,F=443.86,P<0.001;两者交互作用,F=7.89,P=0.041  1 2组人微血管内皮细胞1划痕后各时间点剩余划痕面积 倒置相差显微镜×200,图中标尺为100 μm。1A、1B、1C.分别为P311腺病毒组划痕后0(即刻)、6、11 h,随着划痕后时间的延长,剩余划痕面积逐渐减少,至划痕后11 h划痕基本愈合;1D、1E、1F.分别为空载腺病毒组划痕后0、6、11 h,图1E、1F剩余划痕面积分别较图1B、1C明显增大
1 2组人微血管内皮细胞1划痕后各时间点剩余划痕面积 倒置相差显微镜×200,图中标尺为100 μm。1A、1B、1C.分别为P311腺病毒组划痕后0(即刻)、6、11 h,随着划痕后时间的延长,剩余划痕面积逐渐减少,至划痕后11 h划痕基本愈合;1D、1E、1F.分别为空载腺病毒组划痕后0、6、11 h,图1E、1F剩余划痕面积分别较图1B、1C明显增大2.1.3 细胞血管形成能力
培养8 h,P311腺病毒组细胞管状结构节点数为(731±104)个,明显多于空载腺病毒组的(496±74)个(t=4.49,P=0.001);P311腺病毒组细胞管状结构总长度为(19 231±2 647)μm,明显长于空载腺病毒组[(12 670±2 077)μm,t=4.78,P=0.001]。见图2。
 2 2组人微血管内皮细胞1培养8 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。2A. P311腺病毒组管状结构节点数较多,管状结构总长度较长;2B.空载腺病毒组管状结构节点数少于图2A,管状结构总长度短于图2A
2 2组人微血管内皮细胞1培养8 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。2A. P311腺病毒组管状结构节点数较多,管状结构总长度较长;2B.空载腺病毒组管状结构节点数少于图2A,管状结构总长度短于图2A2.1.4 细胞中VEGFR2/ERK1/2信号通路
转染48 h,与空载腺病毒组比较,P311腺病毒组细胞VEGFR2、ERK1/2蛋白表达量均无明显变化(t值分别为0.28、1.36,P值分别为0.786、0.202),磷酸化VEGFR2、磷酸化ERK1/2蛋白表达量均明显升高(t值分别为17.27、16.08,P<0.001)。见图3。
 3 蛋白质印迹法检测2组人微血管内皮细胞1转染48 h VEGFR2/ERK1/2信号通路相关蛋白的表达量。3A.条带图;3B.条图(样本数为6,
3 蛋白质印迹法检测2组人微血管内皮细胞1转染48 h VEGFR2/ERK1/2信号通路相关蛋白的表达量。3A.条带图;3B.条图(样本数为6,2.2 P311通过VEGFR2/ERK1/2信号通路影响HMEC-1体外血管形成能力
2.2.1 siRNA-VEGFR2对P311高表达引起的VEGFR2/ERK1/2信号通路活化的影响
转染24 h,P311腺病毒+阴性对照siRNA组、空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组和空载腺病毒+siRNA-VEGFR2组细胞磷酸化VEGFR2、VEGFR2和磷酸化ERK1/2蛋白表达量总体比较,差异均有统计学意义(F值分别为27.89、25.07、14.20,P<0.001);4组间细胞ERK1/2蛋白表达量总体比较,差异无统计学意义(F=0.08,P=0.971)。
转染24 h,P311腺病毒+阴性对照siRNA组细胞磷酸化VEGFR2和磷酸化ERK1/2蛋白表达量均明显高于空载腺病毒+阴性对照siRNA组(P值分别为<0.001、0.001),2组细胞VEGFR2和ERK1/2蛋白表达量相近(P值分别为0.063、0.640)。P311腺病毒+阴性对照siRNA组细胞VEGFR2、磷酸化VEGFR2、磷酸化ERK1/2蛋白表达量均明显高于P311腺病毒+siRNA-VEGFR2组(P<0.001),2组细胞ERK1/2蛋白表达量相近(P=0.822)。空载腺病毒+阴性对照siRNA组细胞VEGFR2、磷酸化ERK1/2蛋白表达量均明显高于空载腺病毒+siRNA-VEGFR2组(P值分别为<0.001、0.049),该2组细胞磷酸化VEGFR2和ERK1/2蛋白表达量相近(P值分别为0.071、0.761)。转染24 h,P311腺病毒+siRNA-VEGFR2组和空载腺病毒+siRNA-VEGFR2组细胞VEGFR2、磷酸化VEGFR2、ERK1/2和磷酸化ERK1/2蛋白表达量均相近(P值分别为0.155、0.493、0.952、0.314)。见图4。
 4 蛋白质印迹法检测4组人微血管内皮细胞1转染24 h VEGFR2/ERK1/2信号通路相关蛋白的表达量。4A.条带图;4B.条图(样本数为6,
4 蛋白质印迹法检测4组人微血管内皮细胞1转染24 h VEGFR2/ERK1/2信号通路相关蛋白的表达量。4A.条带图;4B.条图(样本数为6,2.2.2 siRNA-VEGFR2对P311高表达介导的血管形成能力的影响
转染24 h,P311腺病毒+阴性对照siRNA组细胞管状结构节点数明显多于空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组(P值均<0.01),管状结构总长度明显长于空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组(P值均<0.01)。转染24 h,空载腺病毒+阴性对照siRNA组细胞管状结构节点数明显多于空载腺病毒+siRNA-VEGFR2组(P<0.01),管状结构总长度明显长于空载腺病毒+siRNA-VEGFR2组(P<0.01)。P311腺病毒+siRNA-VEGFR2组和空载腺病毒+siRNA-VEGFR2组细胞管状结构节点数和管状结构总长度相近(P>0.05)。见图5、表3。
 5 4组人微血管内皮细胞1转染24 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。5A.P311腺病毒+阴性对照小干扰RNA(siRNA)组管状结构节点数较多,管状结构总长度较长;5B.空载腺病毒+阴性对照siRNA组管状结构节点数明显少于图5A,管状结构总长度明显短于图5A;5C.P311腺病毒+siRNA-血管内皮生长因子受体2(VEGFR2)组管状结构节点数明显少于图5A,管状结构总长度明显短于图5A;5D.空载腺病毒+siRNA-VEGFR2组管状结构节点数明显少于图5B,管状结构总长度明显短于图5B,且均与图5C相近表3 4组人微血管内皮细胞1转染24 h后血管形成情况(
5 4组人微血管内皮细胞1转染24 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。5A.P311腺病毒+阴性对照小干扰RNA(siRNA)组管状结构节点数较多,管状结构总长度较长;5B.空载腺病毒+阴性对照siRNA组管状结构节点数明显少于图5A,管状结构总长度明显短于图5A;5C.P311腺病毒+siRNA-血管内皮生长因子受体2(VEGFR2)组管状结构节点数明显少于图5A,管状结构总长度明显短于图5A;5D.空载腺病毒+siRNA-VEGFR2组管状结构节点数明显少于图5B,管状结构总长度明显短于图5B,且均与图5C相近表3 4组人微血管内皮细胞1转染24 h后血管形成情况(组别 样本数 管状结构节点数(个) 管状结构总长度(μm) P311腺病毒+阴性对照siRNA组 6 720±62 21 241±1 139 空载腺病毒+阴性对照siRNA组 6 428±38 17 005±1 156 P311腺病毒+siRNA- VEGFR2组 6 364±57 13 494±2 465 空载腺病毒+siRNA- VEGFR2组 6 310±75 11 600±2 776 F值 57.07 26.32 P值 <0.001 <0.001 P1值 <0.001 0.002 P2值 <0.001 <0.001 P3值 0.002 <0.001 P4值 0.127 0.121 注:F值、P值为4组间各指标总体比较所得;P1值、P2值分别为P311腺病毒+阴性对照小干扰RNA(siRNA)组与空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-血管内皮生长因子受体2(VEGFR2)组比较所得,P3值、P4值分别为空载腺病毒+siRNA-VEGFR2组与空载腺病毒+阴性对照siRNA组、P311腺病毒+siRNA-VEGFR2组比较所得 2.2.3 ERK1/2抑制剂对P311高表达引起的ERK1/2信号通路活化的影响
处理2 h,P311腺病毒+DMSO组、空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组和空载腺病毒+ERK1/2抑制剂组细胞磷酸化ERK1/2蛋白表达量总体比较,差异有统计学意义(F=16.79,P<0.001);4组细胞ERK1/2 蛋白表达量总体比较,差异无统计学意义(F=0.99,P=0.419)。
P311腺病毒+DMSO组细胞磷酸化ERK1/2蛋白表达量明显高于空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组(P值均<0.001),P311腺病毒+DMSO组与空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组细胞ERK1/2蛋白表达量均相近(P值分别为0.806、0.467)。空载腺病毒+DMSO组细胞磷酸化ERK1/2蛋白表达量明显高于空载腺病毒+ERK1/2抑制剂组(P=0.030),2组细胞ERK1/2 蛋白表达量相近(P=0.140)。P311腺病毒+ERK1/2 抑制剂组和空载腺病毒+ERK1/2抑制剂组细胞磷酸化ERK1/2和ERK1/2蛋白表达量均相近(P值分别为0.591、0.373)。见图6。
 6 蛋白质印迹法检测4组人微血管内皮细胞1处理2 h ERK1/2信号通路相关蛋白的表达量。6A.条带图;6B.条图(样本数为6,
6 蛋白质印迹法检测4组人微血管内皮细胞1处理2 h ERK1/2信号通路相关蛋白的表达量。6A.条带图;6B.条图(样本数为6,2.2.4 ERK1/2抑制剂对P311高表达介导的促HMEC-1体外血管形成作用的影响
于处理2 h,P311腺病毒+DMSO组细胞管状结构节点数明显多于空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组(P值均<0.01),管状结构总长度明显长于空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组(P值均<0.01)。空载腺病毒+DMSO组管状结构节点数明显多于空载腺病毒+ERK1/2抑制剂组(P<0.01),管状结构总长度明显长于空载腺病毒+ERK1/2抑制剂组(P<0.01)。P311腺病毒+ERK1/2 抑制剂组和空载腺病毒+ERK1/2抑制剂组细胞管状结构节点数和总长度均相近(P>0.05)。见图7、表4。
 7 4组人微血管内皮细胞1处理2 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。7A.P311腺病毒+二甲基亚砜(DMSO)组管状结构节点数较多,管状结构总长度较长;7B.空载腺病毒+DMSO组管状结构节点数少于图7A,管状结构总长度短于图7A;7C.P311腺病毒+胞外信号调节激酶1/2(ERK1/2)抑制剂组管状结构节点数少于图7A,管状结构总长度短于图7A;7D.空载腺病毒+ERK1/2抑制剂组管状结构节点数和总长度与图7C相近表4 4组人微血管内皮细胞1处理2 h血管形成情况(
7 4组人微血管内皮细胞1处理2 h血管形成情况 倒置相差显微镜×200,图中标尺为100 μm。7A.P311腺病毒+二甲基亚砜(DMSO)组管状结构节点数较多,管状结构总长度较长;7B.空载腺病毒+DMSO组管状结构节点数少于图7A,管状结构总长度短于图7A;7C.P311腺病毒+胞外信号调节激酶1/2(ERK1/2)抑制剂组管状结构节点数少于图7A,管状结构总长度短于图7A;7D.空载腺病毒+ERK1/2抑制剂组管状结构节点数和总长度与图7C相近表4 4组人微血管内皮细胞1处理2 h血管形成情况(组别 样本数 管状结构节点数(个) 管状结构总长度(μm) P311腺病毒+DMSO组 6 726±72 20 318±1 433 空载腺病毒+DMSO组 6 421±39 16 846±1 464 P311腺病毒+ERK1/2抑制剂组 6 365±41 15 114±1 950 空载腺病毒+ERK1/2抑制剂组 6 317±67 13 188±2 306 F值 63.49 16.53 P值 <0.001 <0.001 P1值 <0.001 0.004 P2值 <0.001 <0.001 P3值 0.005 0.002 P4值 0.162 0.083 注:F值、P值为4组间各指标总体比较所得;DMSO为二甲基亚砜,ERK1/2为胞外信号调节激酶1/2;P1值、P2值分别为P311腺病毒+DMSO组与空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组比较所得,P3值、P4值分别为空载腺病毒+ERK1/2抑制剂组与空载腺病毒+DMSO组、P311腺病毒+ERK1/2抑制剂组比较所得 3. 讨论
创面愈合过程被划分为4个连续而又有重叠的阶段,包括凝血期、炎症期、增殖期和重构期[25]。创面一旦形成,就开始了局部新的血液供应系统的建立,该进程贯穿整个创面修复过程[2],但其机制仍不完全清楚。本课题组前期研究表明,皮肤微血管内皮细胞在静息状态下低表达P311,在炎症刺激条件下高表达P311,进而参与创面血管新生[22]。本研究进一步证实,P311能够通过激活VEGFR2/ERK1/2 信号通路,发挥促进HMEC-1血管形成的作用。
P311基因是一种损伤环境诱导型基因,即在静息状态下低表达或者不表达,当受到外界环境刺激时表达升高,从而发挥相关生物学功能[22, 23]。该基因能够编码一种种属间高度保守的细胞内多功能蛋白[10]。在创面愈合过程中,P311持续高表达,可增强表皮干细胞Rho A和Rac1蛋白的活性[21],并激活TGF-β1/Smad信号通路[23],从而发挥促进创面修复作用。近年来研究表明,P311表达于创面皮肤微血管内皮细胞,P311基因敲除导致皮肤创面愈合过程中的血管新生减弱,进而导致创面愈合延迟;P311基因敲除皮肤微血管内皮细胞迁移及血管形成能力均减弱[22]。这些研究结果提示P311可能通过影响微血管内皮细胞的迁移和血管形成能力参与血管新生。微血管内皮细胞具有正常有效的增殖能力在血管形成中也至关重要[26]。
本研究用P311腺病毒转染HMEC-1,CCK-8法检测结果显示,P311腺病毒组和空载腺病毒组HMEC-1增殖能力没有明显差异;划痕试验结果显示,P311腺病毒组HMEC-1迁移能力较空载腺病毒组明显增强;体外血管形成实验结果显示,P311腺病毒组HMEC-1血管形成能力较空载腺病毒组明显增强。以上结果表明P311高表达能够显著提高HMEC-1的迁移能力和体外血管形成能力,而对HMEC-1增殖能力没有明显影响。
VEGFR2信号通路在启动血管内皮细胞各项生物学过程中发挥重要作用[27]。VEGFR2活化是VEGFR2同二聚化或异二聚化导致酪氨酸激酶的活化和受体细胞内结构域中酪氨酸残基的自磷酸化[28, 29]。VEGFR2活化后启动血管内皮细胞内多种信号通路[30]。研究表明,在小鼠中VEGFR2 Tyr1173磷酸化(人类中Tyr1175磷酸化)对ERK1/2 激活至关重要[31]。ERK1/2信号通路与细胞的成活[32]、迁移[33]、血管形成[34]等生物学过程密切相关[35]。然而,此前尚鲜见有关P311对VEGFR2活化和ERK1/2活化的影响的研究。本研究采用蛋白质印迹法检测P311腺病毒组和空载腺病毒组HMEC-1中VEGFR2、磷酸化VEGFR2、ERK1/2和磷酸化ERK1/2的蛋白表达水平,结果显示,P311高表达对HMEC-1 VEGFR2、ERK1/2的蛋白表达没有明显影响,但是却能够促进VEGFR2、ERK1/2的磷酸化。根据上述结果可以得出,P311高表达可以激活VEGFR2/ERK1/2信号通路,该机制可能参与了P311促HMEC-1体外血管形成的作用。
为进一步探讨P311介导的促体外血管形成作用是否通过激活VEGFR2/ERK1/2信号通路而发挥作用,本研究分别采用siRNA-VEGFR2和ERK抑制剂SCH772984阻断VEGFR2/ERK1/2信号通路的活化,随后采用蛋白质印迹法检测细胞VEGFR2磷酸化和ERK1/2磷酸化水平的变化,采用体外血管形成实验检测细胞血管形成能力的变化。结果显示,P311腺病毒+阴性对照siRNA组细胞磷酸化VEGFR2和磷酸化ERK1/2蛋白表达水平均明显高于空载腺病毒+阴性对照siRNA组,提示P311高表达能够促进VEGFR2/ERK1/2信号通路活化。在阻断VEGFR2途径后,VEGFR2和ERK1/2磷酸化水平均明显降低,HMEC-1体外血管形成能力明显减弱,提示siRNA-VEGFR2能够抑制P311高表达引起的VEGFR2/ERK1/2信号通路活化,进而抑制P311高表达介导的促HMEC-1体外血管形成作用。另外,在抑制ERK1/2信号通路后,ERK1/2磷酸化水平均明显降低,HMEC-1体外血管形成能力明显减弱,提示ERK1/2抑制剂能够抑制P311高表达引起的ERK1/2信号通路活化,进而抑制P311高表达介导的促HMEC-1体外血管形成作用。由此推测,P311通过激活VEGFR2/ERK1/2信号通路发挥促进HMEC-1血管形成的作用。
综上所述,P311在创面血管新生中发挥重要作用,能够通过激活VEGFR2/ERK1/2信号通路发挥促进HMEC-1血管形成的作用。本研究结果丰富了创面愈合理论,为促进皮肤创面修复提供了新思路、新线索与新靶点。
·《Burns & Trauma》好文推荐·
严重烫伤后肠道菌群移位的分子机制:囊性纤维化穿膜传导调节蛋白的作用
陈文涛:研究设计、数据分析、文章的撰写及修改;王小祥:数据分析、图表绘制、文章修改;郑文炼、张伟强、毛璐佳、卓佳楠:数据采集与数据分析;周思彤:研究指导与数据采集;杨荣华:研究设计、研究指导、文章修改和经费支持所有作者均声明不存在利益冲突 -
参考文献
(30) [1] WangZC, ZhaoWY, CaoY, et al. The roles of inflammation in keloid and hypertrophic scars[J]. Front Immunol, 2020,11:603187. DOI: 10.3389/fimmu.2020.603187. [2] ChiangRS, BorovikovaAA, KingK, et al. Current concepts related to hypertrophic scarring in burn injuries[J]. Wound Repair Regen, 2016,24(3):466-477. DOI: 10.1111/wrr.12432. [3] LeeHJ, JangYJ. Recent understandings of biology, prophylaxis and treatment strategies for hypertrophic scars and keloids[J]. Int J Mol Sci, 2018,19(3):711.DOI: 10.3390/ijms19030711. [4] 郑建新,张亚军. 二氧化碳点阵激光联合曲安奈德注射治疗增生性瘢痕的疗效[J]. 中国医疗美容,2020,10(5):84-88. DOI: 10.19593/j.issn.2095-0721.2020.05.020. [5] NiuM, ChenP. Crosstalk between gut microbiota and sepsis[J/OL]. Burns Trauma, 2021,9:tkab036[2023-11-29]. https://pubmed.ncbi.nlm.nih.gov/34712743/. DOI: 10.1093/burnst/tkab036. [6] CamposLF, TagliariE, CasagrandeT, et al. Effects of probiotics supplementation on skin wound healing in diabetic rats[J]. Arq Bras Cir Dig, 2020,33(1):e1498. DOI: 10.1590/0102-672020190001e1498. [7] AlamA, NeishA. Role of gut microbiota in intestinal wound healing and barrier function[J]. Tissue Barriers, 2018,6(3):1539595. DOI: 10.1080/21688370.2018.1539595. [8] 吴金春, 刘彦民, 苏晓灵. 肠道菌群/肠道微生态与心血管疾病发生的关系研究进展[J].解放军医学杂志,2023,48(7):851-855. DOI: 10.11855/j.issn.0577-7402.2696.2022.0830. [9] WaasdorpM, KromBP, BikkerFJ, et al. The bigger picture: why oral mucosa heals better than skin[J]. Biomolecules, 2021,11(8):1165.DOI: 10.3390/biom11081165. [10] PatelBK, PatelKH, HuangRY, et al. The gut-skin microbiota axis and its role in diabetic wound healing-a review based on current literature[J]. Int J Mol Sci, 2022, 23(4):2375. DOI: 10.3390/ijms23042375. [11] BurgessS, FoleyCN, AllaraE, et al. A robust and efficient method for Mendelian randomization with hundreds of genetic variants[J]. Nat Commun, 2020,11(1):376. DOI: 10.1038/s41467-019-14156-4. [12] LevinMG, JudyR, GillD, et al. Genetics of height and risk of atrial fibrillation: a Mendelian randomization study[J]. PLoS Med, 2020,17(10):e1003288. DOI: 10.1371/journal.pmed.1003288. [13] O'NeillCA, MonteleoneG, McLaughlinJT, et al. The gut-skin axis in health and disease: a paradigm with therapeutic implications[J]. Bioessays, 2016,38(11):1167-1176. DOI: 10.1002/bies.201600008. [14] BelkaidY, HarrisonOJ. Homeostatic immunity and the microbiota[J]. Immunity, 2017,46(4):562-576. DOI: 10.1016/j.immuni.2017.04.008. [15] Polkowska-PruszyńskaB, GerkowiczA, KrasowskaD. The gut microbiome alterations in allergic and inflammatory skin diseases - an update[J]. J Eur Acad Dermatol Venereol, 2020,34(3):455-464. DOI: 10.1111/jdv.15951. [16] BenyacoubJ, BoscoN, BlanchardC, et al. Immune modulation property of Lactobacillus paracasei NCC2461 (ST11) strain and impact on skin defences[J]. Benef Microbes, 2014,5(2):129-136. DOI: 10.3920/BM2013.0014. [17] PeralMC, RachidMM, GobbatoNM, et al. Interleukin-8 production by polymorphonuclear leukocytes from patients with chronic infected leg ulcers treated with Lactobacillus plantarum[J]. Clin Microbiol Infect, 2010,16(3):281-286. DOI: 10.1111/j.1469-0691.2009.02793.x. [18] AfoudaP, DurandGA, LagierJC, et al. Noncontiguous finished genome sequence and description of Intestinimonas massiliensis sp. nov strain GD2T, the second Intestinimonas species cultured from the human gut[J]. Microbiologyopen, 2019,8(1):e00621. DOI: 10.1002/mbo3.621. [19] ZhouJ, LiM, ChenQ, et al. Programmable probiotics modulate inflammation and gut microbiota for inflammatory bowel disease treatment after effective oral delivery[J]. Nat Commun, 2022,13(1):3432. DOI: 10.1038/s41467-022-31171-0. [20] LiuJ, ChangG, HuangJ, et al. Sodium butyrate inhibits the inflammation of lipopolysaccharide-induced acute lung injury in mice by regulating the Toll-like receptor 4/nuclear factor κB signaling pathway[J]. J Agric Food Chem, 2019,67(6):1674-1682. DOI: 10.1021/acs.jafc.8b06359. [21] ArpaiaN, CampbellC, FanX, et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation[J]. Nature, 2013,504(7480):451-455. DOI: 10.1038/nature12726. [22] HuangC, AkaishiS, HyakusokuH, et al. Are keloid and hypertrophic scar different forms of the same disorder? A fibroproliferative skin disorder hypothesis based on keloid findings[J]. Int Wound J, 2014,11(5):517-522. DOI: 10.1111/j.1742-481X.2012.01118.x. [23] HuangC, OgawaR. Role of inflammasomes in keloids and hypertrophic scars-lessons learned from chronic diabetic wounds and skin fibrosis[J]. Int J Mol Sci, 2022,23(12):6280.DOI: 10.3390/ijms23126820. [24] ZhangT, WangXF, WangZC, et al. Current potential therapeutic strategies targeting the TGF-β/Smad signaling pathway to attenuate keloid and hypertrophic scar formation[J]. Biomed Pharmacother, 2020,129:110287. DOI: 10.1016/j.biopha.2020.110287. [25] La ReauAJ, SuenG. The Ruminococci: key symbionts of the gut ecosystem[J]. J Microbiol, 2018,56(3):199-208. DOI: 10.1007/s12275-018-8024-4. [26] LiY, ZhangSX, YinXF, et al. The gut microbiota and its relevance to peripheral lymphocyte subpopulations and cytokines in patients with rheumatoid arthritis[J]. J Immunol Res, 2021,2021:6665563. DOI: 10.1155/2021/6665563. [27] ZhouN, ShenY, FanL, et al. The characteristics of intestinal-barrier damage in rats with IgA nephropathy[J]. Am J Med Sci, 2020,359(3):168-176. DOI: 10.1016/j.amjms.2019.11.011. [28] WangT, SternesPR, GuoXK, et al. Autoimmune diseases exhibit shared alterations in the gut microbiota[J]. Rheumatology (Oxford), 2024,63(3):856-865. DOI: 10.1093/rheumatology/kead364. [29] Lopez-SilesM, KhanTM, DuncanSH, et al. Cultured representatives of two major phylogroups of human colonic Faecalibacterium prausnitzii can utilize pectin, uronic acids, and host-derived substrates for growth[J]. Appl Environ Microbiol, 2012,78(2):420-428. DOI: 10.1128/AEM.06858-11. [30] MahmudMR, AkterS, TamannaSK, et al. Impact of gut microbiome on skin health: gut-skin axis observed through the lenses of therapeutics and skin diseases[J]. Gut Microbes, 2022,14(1):2096995. DOI: 10.1080/19490976.2022.2096995. -
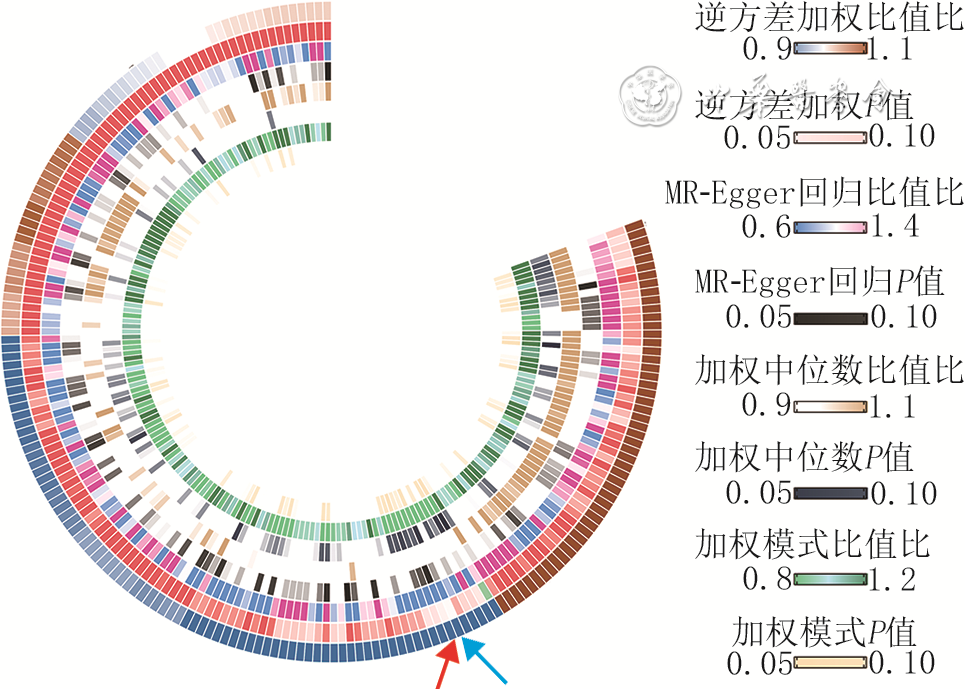
2 人的196个肠道菌群单核苷酸多态性与增生性瘢痕因果关系的双样本孟德尔随机化分析
注:从外到内的圆圈依次代表肠道菌群的逆方差加权的比值比和P值、MR-Egger回归的比值比和P值、加权中位数的比值比和P值、加权模式的比值比和P值,同一个圆心的不同扇环块均代表一种肠道细菌,外圈中蓝色箭头指示肠杆菌属、红色箭头指示瘤胃球菌2属
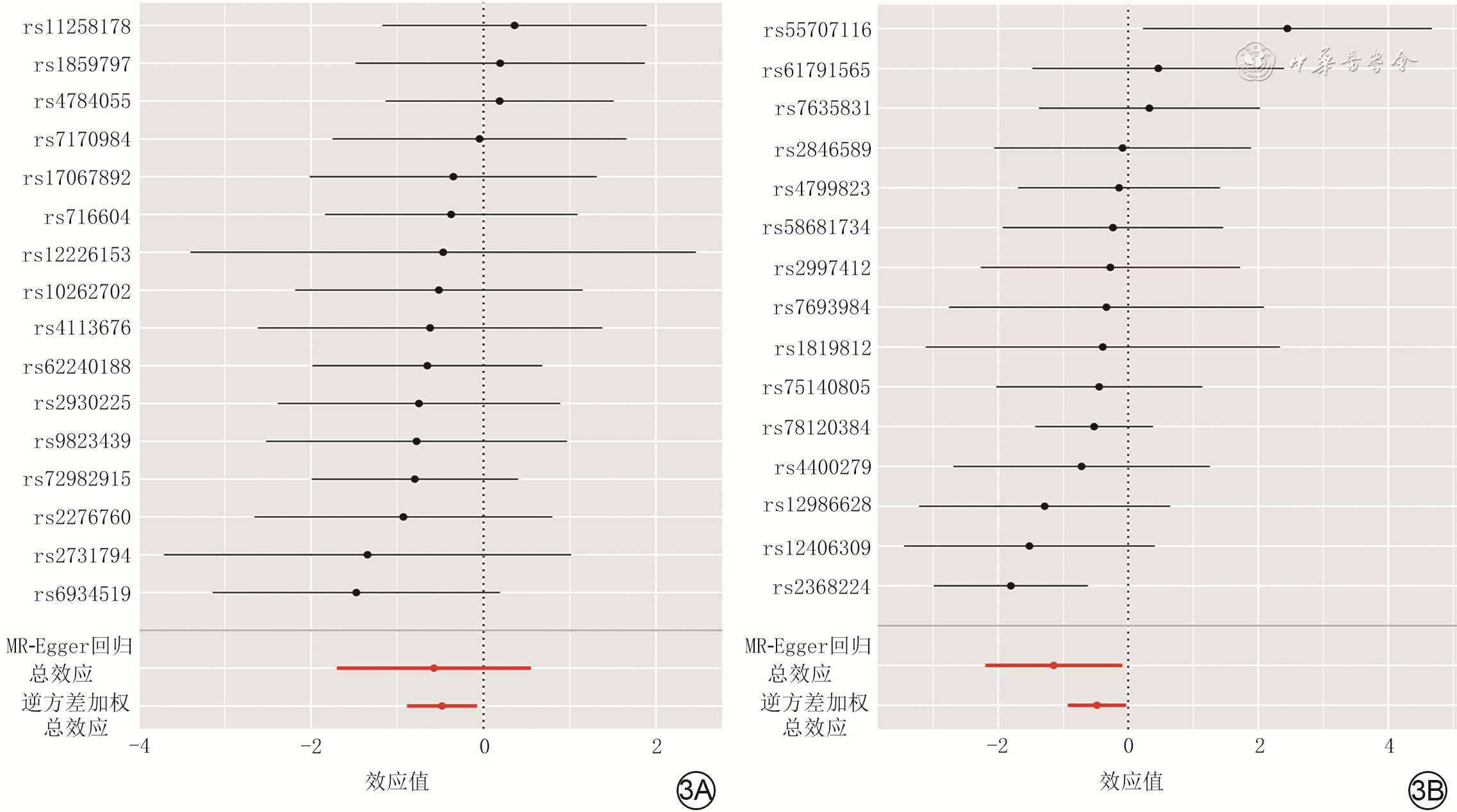
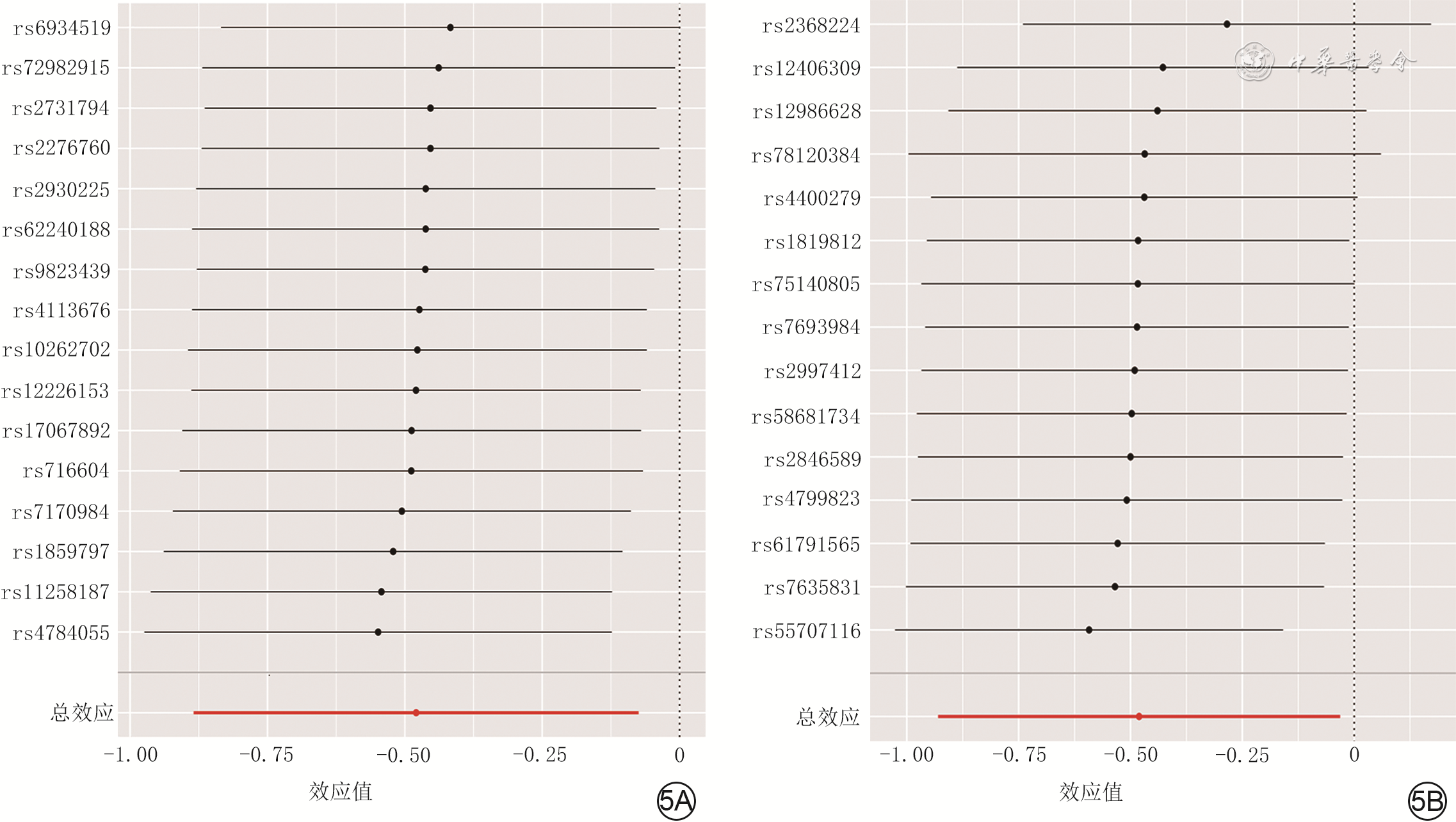
3 逆方差加权和MR-Egger回归分析人肠杆菌属16个SNP和瘤胃球菌2属15个SNP分别与增生性瘢痕因果关系的森林图。3A.肠杆菌属;3B.瘤胃球菌2属
注:线段中的圆点为比值比,该值>0为危险因素、<0为保护因素;SNP为单核苷酸多态性
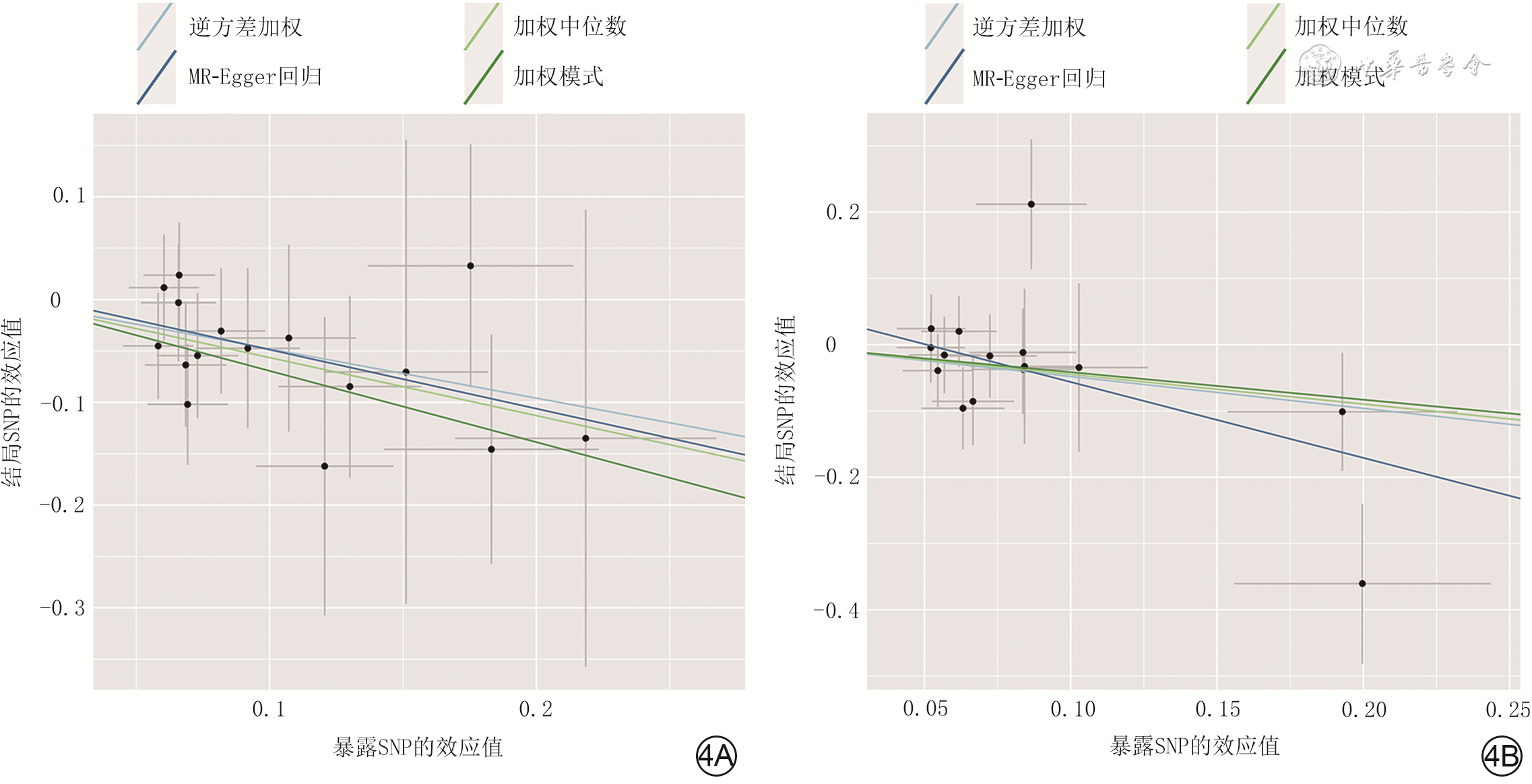
4 双样本孟德尔随机化分析人肠杆菌属16个SNP和瘤胃球菌2属15个SNP分别与增生性瘢痕之间因果关系的散布图。4A.肠杆菌属;4B.瘤胃球菌2属
注:SNP为单核苷酸多态性;直线的斜率表示每种方法的因果关联,斜率为负值表明肠道菌群SNP可降低增生性瘢痕发生风险
-

 下载:
下载:
 下载:
下载:

计量
- 文章访问数: 3677
- HTML全文浏览量: 163
- PDF下载量: 47
- 被引次数: 0
