



Analysis of the types and functions of CD34 + cells in full-thickness skin defect wounds of normal mice and diabetic mice by single-cell RNA sequencing
-
摘要:
目的 单细胞RNA测序解析普通小鼠和糖尿病小鼠全层皮肤缺损创面中CD34 +细胞的类型与功能。 方法 该研究为实验研究。构建CD34 +细胞谱系追踪小鼠,实现CD34 +细胞在荧光条件下可视化。取6只7~8周龄雄性CD34 +细胞谱系追踪小鼠(设为糖尿病组),腹腔注射链脲佐菌素构建糖尿病模型,于小鼠13周龄时在其背部制备全层皮肤缺损创面;另取6只13周龄雄性CD34 +细胞谱系追踪小鼠(设为对照组),在其背部制备全层皮肤缺损创面。伤后4 d,分别收集对照组3只小鼠和糖尿病组2只小鼠创面组织,消化制备单细胞悬液,采用荧光活化细胞分选法筛选出CD34 +细胞后进行单细胞RNA测序,采用R语言的Seurat 4.0.2程序通过降维可视化和细胞聚类分析CD34 +细胞类型并筛选注释各CD34 +细胞亚群的标记基因,对2组小鼠创面组织间CD34 +成纤维细胞(Fb)、平滑肌细胞、角质形成细胞(KC)、类软骨细胞的差异表达基因(DEG)进行京都基因与基因组百科全书(KEGG)和基因本体论(GO)富集分析,探索细胞功能。 结果 伤后4 d,2组小鼠创面组织中CD34 +细胞均包含7种细胞类型,具体为内皮细胞、Fb、KC、巨噬细胞、T细胞、平滑肌细胞和类软骨细胞,其中Fb细分为5个亚群。与对照组比较,糖尿病组小鼠创面组织中的CD34 +内皮细胞、Fb亚群1、Fb亚群4、KC、类软骨细胞占比升高,CD34 +Fb亚群2、Fb亚群3和平滑肌细胞占比下降。注释CD34 +类软骨细胞、内皮细胞、Fb亚群1、Fb亚群2、Fb亚群3、Fb亚群4、Fb亚群5、KC、巨噬细胞、平滑肌细胞、T细胞的标记基因分别为转移相关肺腺癌转录本1、脂肪酸结合蛋白4、Gremlin 1、补体成分4B、H19印记母源表达转录本、Dickkopf Wnt信号通路抑制剂2、纤维调节蛋白、角蛋白5、CD74分子、G蛋白信号调节蛋白5、可诱导T细胞共刺激分子。KEGG和GO富集分析显示,与对照组比较,糖尿病组小鼠伤后4 d创面组织中CD34 +Fb差异表达显著的DEG显著富集于炎症反应、细胞外基质(ECM)组装、细胞增殖调控、衰老相关条目( P值均<0.05),CD34 +平滑肌细胞差异表达显著的DEG显著富集于细胞迁移、凋亡过程、转录的正调控、吞噬体等条目( P值均<0.05),CD34 +KC差异表达显著的DEG显著富集于线粒体功能、转录、神经退行性疾病相关条目( P值均<0.05),CD34 +类软骨细胞差异表达显著的DEG显著富集于节律调控、ECM、病毒感染等相关条目( P值均<0.05)。 结论 CD34 +细胞在普通小鼠和糖尿病小鼠全层皮肤缺损创面愈合过程中均存在高异质性,2种小鼠创面间CD34 +细胞亚群差异表达显著的DEG显著富集的相关功能与创面愈合过程紧密相关。 Abstract:Objective To analyze the types and functions of CD34 + cells in full-thickness skin defect wounds of normal mice and diabetic mice by single-cell RNA sequencing. Methods This study was an experimental study. The CD34 + cell lineage tracing mouse was produced, and the visualization of CD34 + cells under the fluorescent condition was realized. Six male CD34 + cell lineage tracing mice aged 7-8 weeks (designated as diabetic group) were intraperitoneally injected with streptozotocin to establish a diabetic model, and full-thickness skin defect wounds were prepared on their backs when they reached 13 weeks old. Another 6 male CD34 + cell lineage tracing mice aged 13 weeks (designated as control group) were also subjected to full-thickness skin defect wounds on their backs. On post-injury day (PID) 4, wound tissue was collected from 3 mice in control group and 2 mice in diabetic group, and digested to prepare single-cell suspensions. CD34 + cells were screened using fluorescence-activated cell sorting, followed by single-cell RNA sequencing. The Seurat 4.0.2 program in the R programming language was utilized for dimensionality reduction, visualization, and cell clustering analysis of CD34 + cell types, and to screen and annotate the marker genes for each CD34 + cell subpopulation. Kyoto encyclopedia of genes and genomes (KEGG) and gene ontology (GO) enrichment analysis was performed to analyze the differentially expressed genes (DEGs) of CD34 + fibroblasts (Fbs), smooth muscle cells (SMCs), keratinocytes (KCs), and chondrocyte-like cells (CLCs) in the wound tissue of two groups of mice for exploring cellular functions. Results On PID 4, CD34 + cells in the wound tissue of both groups of mice were consisted of 7 cell types, specifically endothelial cells, Fbs, KCs, macrophages, T cells, SMCs, and CLCs. Among these, Fbs were further classified into 5 subpopulations. Compared with those in control group, the proportions of CD34 + endothelial cells, Fbs subpopulation 1, Fbs subpopulation 4, KCs, and CLCs in the wound tissue of mice were increased in diabetic group, while the proportions of CD34 + Fbs subpopulation 2, Fbs subpopulation 3, and SMCs were decreased. The marker genes for annotating CD34 + CLCs, endothelial cells, Fbs subpopulation 1, Fbs subpopulation 2, Fbs subpopulation 3, Fbs subpopulation 4, Fbs subpopulation 5, KCs, macrophages, SMCs, and T cells were respectively metastasis-associated lung adenocarcinoma transcript 1, fatty acid binding protein 4, Gremlin 1, complement component 4B, H19 imprinted maternally expressed transcript, Dickkopf Wnt signaling pathway inhibitor 2, fibromodulin, keratin 5, CD74 molecule, regulator of G protein signaling 5, and inducible T-cell co-stimulator molecule. KEGG and GO enrichment analysis revealed that, compared with those in control group, DEGs with significant differential expression (SDE) in CD34 + Fbs from the wound tissue of mice in diabetic group on PID 4 were significantly enriched in terms related to inflammatory response, extracellular matrix (ECM) organization, regulation of cell proliferation, and aging (with Pvalues all <0.05), DEGs with SDE in CD34 + SMCs were significantly enriched in terms related to cell migration, apoptotic process, positive regulation of transcription, and phagosome (with P values all <0.05), DEGs with SDE in CD34 + KCs were significantly enriched in terms related to mitochondrial function, transcription, and neurodegenerative diseases (with P values all <0.05), and DEGs with SDE in CD34 + CLCs were significantly enriched in terms related to rhythm regulation, ECM, and viral infection (with P values all <0.05). Conclusions CD34 + cells display high heterogeneity in the healing process of full-thickness skin defect wounds in both normal mice and diabetic mice. The significantly enriched functions of DEGs with SDE in CD34 + cell subpopulations in the wound tissue of the two mouse groups are closely related to the wound healing process. -
Key words:
- Wound healing /
- Skin /
- CD34 /
- Single-cell RNA sequencing /
- Cell subpopulations /
- Marker genes /
- Cellular heterogeneity /
- Cell function
-
烧伤是常见的损伤之一,我国每年有数十万人因为烧伤而致残或致畸。烧伤后病理性瘢痕组织的形成是烧伤患者康复过程中的重要并发症之一,不仅严重影响患者的外观,而且可引起器官及组织的功能障碍,严重影响患者生活质量及身心健康[1]。对于瘢痕产生的机制,目前在微观方面的研究仅涉及细胞因子、ECM等,而其分子机制尚不十分明确[2]。
有研究显示,青光眼滤过术后使用表观遗传修饰可减少瘢痕组织的产生[3],表明瘢痕产生的过程中可能存在表观遗传改变。DNA甲基化是一种重要的表观遗传修饰机制,可在不改变DNA序列的情况下改变基因表达量[4, 5]。甲基化水平的高低会显著影响蛋白的最终翻译。生物信息学分析是一项新兴的研究方法,可结合测序数据进行全基因组分析,有针对性地寻找与疾病相关的基因或基因集[6, 7]。甲基化研究在瘢痕中较为常见,但检索显示以生物信息学为基础,探究甲基化与瘢痕形成关系的研究罕见,因此以生物信息学为基础探讨甲基化与瘢痕形成的关系显得十分必要。
加权基因共表达网络分析(weighted gene co-expression network analysis,WGCNA)可对大量芯片来源的基因数据进行有效分析,是生物信息学研究的重要工具[8]。本观察性研究使用WGCNA对正常组织和瘢痕组织的mRNA表达谱进行分析并结合甲基化数据集对选定模块内的基因进行甲基化状态判断,对烧伤后瘢痕组织形成的分子机制进行探讨,并使用支持向量机(support vector machine,SVM)模型进行进一步验证,为临床治疗烧伤后瘢痕组织提供新思路。
1. 资料及方法
1.1 数据收集
从美国国家生物技术信息中心基因表达综合数据库(
https://www.ncbi.nlm.nih.gov/geo/ )内检索并下载相关数据集,即检索基因表达数据库(gene expression omnibus,GEO)中烧伤后瘢痕组织相关数据集,结果显示mRNA数据集GSE136906及甲基化数据集GSE137134是同一批次的12个样本(均由澳大利亚大学生物医学院烧伤研究组上传,每个数据集均为6个样本),分别进行了mRNA测序和甲基化测序,因此纳入本研究;数据集GSE108110在剩余数据集中样本量相对较大,因此纳入SVM及建模分析(表1)。表1 烧伤后瘢痕组织相关数据集的详细信息数据集 测序平台 Affymetrix eneChip文件 样本数(个) 瘢痕数(个) GSE137134 GPL13534-11288 1 6 6 GSE136906 GPL16686 2 6 6 GSE108110 GPL570 3 9 9 注:1、2、3分别为Illumina HumanMethylation450 BeadChip (HumanMethylation450_15017482)、[HuGene-2_0-st] Affymetrix Human Gene 2.0 ST Array [transcript (gene) version]、[HG-U133_Plus_2] Affymetrix Human Genome U133 Plus 2.0 Array 1.2 GEO数据集的分析
下载GSE137134与GSE136906数据集并将数据集中的探针代码转换成基因符号。使用R语言中的“Limma”软件包寻找GSE137134中瘢痕组织与正常组织的差异甲基化基因(differentially methylated genes,DMG)和GSE136906中的差异表达基因(differently expressed genes,DEG),即寻找P<0.05相关基因并记录,将这些基因纳入WGCNA分析。
1.3 WGCNA构建及临床相关性分析
使用R语言软件中的“FlashClust”软件包对样本进行聚类分析。使用“pickSoftThreshold”函数来调节参数β的权重以尽量符合无尺度网络。使用WGCNA软件包将有相关性和相邻关系的DMG计算成为拓扑重叠矩阵(opological overlap matrix, TOM)并计算其相应的相异度1-TOM。使用1-TOM作为距离度量,进行分层聚类以识别模块。设定模块内最少的基因个数为30个,将高度相似的模块通过聚类标记并合并。使用“plotDendroAndColors”函数对基因模块进行可视化,并选取模块内基因绘制热图。将临床特征和模块内的基因联合分析,寻找与瘢痕组织密切相关的模块并探究模块内基因的生物学意义,对生成模块与临床特征之间的相关性热图进行聚类。
1.4 模块内基因功能富集分析
使用Metascape数据库和基因探针富集分析(gene set enrichment analysis,GSEA)软件对模块内的基因进行基因本体论(gene ontology,GO)分析和京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)富集分析和注释。
1.5 模块内基因甲基化状态的探究
基于瘢痕组织mRNA数据集GSE136906及其甲基化数据集GSE137134属于同一批次的样本,本研究利用数据集GSE136906探究模块内基因的甲基化状态。使用Funrich软件将模块内高甲基化状态的基因与低表达的基因取交集,将模块内低甲基化状态的基因与高表达的基因取交集,在交集内的基因是具有异常甲基化状态的基因。
1.6 统计学处理
依据受试者操作特征(ROC)曲线下面积,评估基因的诊断价值。使用数据集GSE136906及GSE108110进行机器学习,将这2个数据集内异常甲基化基因的表达量依次纳入SVM模型并进行标准化,构建SVM分类器,使用交叉验证的方法选取惩罚参数c和核函数参数g,随机选取2/3的样本作为训练集,其余1/3则作为验证集,当SVM分类器的效能达到90%时则不再纳入新基因。
使用SPSS 22.0、GraphPad Prism 8统计软件进行数据处理及统计学分析,使用Matlab软件进行支持向量机模型构建,P<0.05为差异有统计学意义。
2. 结果
2.1 WGCNA网络构建及临床相关性分析
从数据集GSE136906中筛选出1 061个DEG,见图1。从数据集GSE137134中筛选出16 141个DMG。聚类结果显示没有明显离群的样本,因此2个数据集中12个样本均被纳入共表达网络。“pickSoftThreshold”函数的结果显示,当权重参数β=9时,log(k)与log [p(k)]之间相关系数的平方>0.9,k表示节点的连接度,p(k)表示WGCNA网络具有无标度网络的特征。选取软阈值β=9来构建WGCNA网络(图2)。模块的相关性热图与聚类分析展示在图3中。筛选后得到10个相应的模块,临床相关性结果显示棕色模块与瘢痕组织形成相关性较高(图4),该模块内共有2 646个基因。在所有相关系数>0.7的模块中,棕色模块内基因数目最多,因此选择该模块进行进一步研究。
 1 GSE136906数据集中烧伤后瘢痕和正常组织的差异表达基因火山图注: P-value为差异表达分析后未经校正的P值,Fold change为瘢痕组织与正常组织差异表达倍数;图中绿色为低表达且差异有统计学意义的基因,红色为高表达且差异有统计学意义的基因,黑色为差异无统计学意义的基因
1 GSE136906数据集中烧伤后瘢痕和正常组织的差异表达基因火山图注: P-value为差异表达分析后未经校正的P值,Fold change为瘢痕组织与正常组织差异表达倍数;图中绿色为低表达且差异有统计学意义的基因,红色为高表达且差异有统计学意义的基因,黑色为差异无统计学意义的基因 3 加权基因共表达网络分析网络模块之间的相互作用注:MEgreen、MEbrown、MEturquoise、MEmagenta、MEpink、MEblack、MEyellow、MEblue、MEred分别表示绿、棕、蓝绿、洋红、粉、黑、黄、蓝、红色模块;图中横向刻度值为各模块聚类分析相对聚类距离;图中纵向刻度值为数据模块色值
3 加权基因共表达网络分析网络模块之间的相互作用注:MEgreen、MEbrown、MEturquoise、MEmagenta、MEpink、MEblack、MEyellow、MEblue、MEred分别表示绿、棕、蓝绿、洋红、粉、黑、黄、蓝、红色模块;图中横向刻度值为各模块聚类分析相对聚类距离;图中纵向刻度值为数据模块色值 4 烧伤后瘢痕组织临床特征与模块之间的关联注:图左侧标注为加权基因共表达网络分析结果所示颜色模块名称;模块中数据表示对应加权基因共表达网络分析模块与临床性状的相关性系数以及其对应P值,括号内为P值
4 烧伤后瘢痕组织临床特征与模块之间的关联注:图左侧标注为加权基因共表达网络分析结果所示颜色模块名称;模块中数据表示对应加权基因共表达网络分析模块与临床性状的相关性系数以及其对应P值,括号内为P值2.2 棕色模块中基因功能富集分析
GO分析结果显示,模块内的基因主要富集在“α-氨基-3-羟基-5-甲基-4-异恶唑-丙酸酯选择性谷氨酸受体活性的调控”“雄激素受体信号通路的调控”等,见图5A。KEGG通路富集分析结果显示,模块内的基因主要与“细胞因子-细胞因子受体相互作用”“过氧化物酶体增殖物激活受体信号通路”等相关,见图5B。Metascape分析结果显示,模块内基因主要富集在“急性粒细胞白血病”“半胱氨酸型内肽酶抑制剂的活性”“胰岛素分泌的正调节”等方面,见图6。富集分析结果表明,这些基因可通过细胞因子受体、胰岛素受体等调节瘢痕组织。
 5 对烧伤后瘢痕组织相关性高的模块通过基因探针富集分析进行基因功能和通路分析。5A.基因本体论功能富集;5B.京都基因与基因组百科全书功能富集注:图5A中的绿、棕、蓝、黑、浅棕、紫色曲线分别表示对核因子κB导入细胞核的负向调节、对白细胞介素1产生的正向调节、对α-氨基-3-羟基-5-甲基-4-异恶唑-丙酸酯选择性谷氨酸受体的活性调控、雄激素受体信号通路调控、脂肪酸氧化的调控、泛酸样蛋白结合酶的调节通路,图5B中的绿、棕、蓝、黑、浅棕色曲线分别表示细胞因子-细胞因子受体的相互作用、Janus激酶/信号转导与转录激活子信号通路、泛酸和辅酶A生物合成、过氧化物酶体增殖物激活受体信号途径、转化生长因子β信号途径
5 对烧伤后瘢痕组织相关性高的模块通过基因探针富集分析进行基因功能和通路分析。5A.基因本体论功能富集;5B.京都基因与基因组百科全书功能富集注:图5A中的绿、棕、蓝、黑、浅棕、紫色曲线分别表示对核因子κB导入细胞核的负向调节、对白细胞介素1产生的正向调节、对α-氨基-3-羟基-5-甲基-4-异恶唑-丙酸酯选择性谷氨酸受体的活性调控、雄激素受体信号通路调控、脂肪酸氧化的调控、泛酸样蛋白结合酶的调节通路,图5B中的绿、棕、蓝、黑、浅棕色曲线分别表示细胞因子-细胞因子受体的相互作用、Janus激酶/信号转导与转录激活子信号通路、泛酸和辅酶A生物合成、过氧化物酶体增殖物激活受体信号途径、转化生长因子β信号途径 6 对烧伤后瘢痕组织相关性高的模块通过Metascape数据库进行基因功能和通路分析。6A.基因本体论功能富集;6B. 京都基因与基因组百科全书功能富集注:P-value为差异表达分析后未经校正的P值;图6A的红、蓝、绿、紫、橙、黄、棕、浅紫、灰、浅绿、浅黄、浅紫、浅橙色圆点分别与图6B从上至下的条带一一对应,依次表示急性粒细胞白血病、半胱氨酸型内肽酶抑制剂的活性、胰岛素分泌的正调节、突触膜、脂肪细胞分化、高尔基体膜、参与免疫反应的水解酶激活、溶酶体膜、参与免疫反应的白细胞激活、核包膜、神经发生正向调节、中毒反应、心脏发育通路
6 对烧伤后瘢痕组织相关性高的模块通过Metascape数据库进行基因功能和通路分析。6A.基因本体论功能富集;6B. 京都基因与基因组百科全书功能富集注:P-value为差异表达分析后未经校正的P值;图6A的红、蓝、绿、紫、橙、黄、棕、浅紫、灰、浅绿、浅黄、浅紫、浅橙色圆点分别与图6B从上至下的条带一一对应,依次表示急性粒细胞白血病、半胱氨酸型内肽酶抑制剂的活性、胰岛素分泌的正调节、突触膜、脂肪细胞分化、高尔基体膜、参与免疫反应的水解酶激活、溶酶体膜、参与免疫反应的白细胞激活、核包膜、神经发生正向调节、中毒反应、心脏发育通路2.3 棕色模块内基因甲基化状态
共有35个具有异常甲基化状态的基因,其中的高表达-低甲基化基因21个(图7A、7B),低表达-高甲基化基因14个(图7C、7D)。
 7 烧伤后瘢痕组织差异表达甲基化基因的热图。7A.21个高表达-低甲基化基因的热图;7B.14个低表达-高甲基化基因的热图注:“type”表示组织类型,“N”表示正常组织,“S”表示瘢痕组织
7 烧伤后瘢痕组织差异表达甲基化基因的热图。7A.21个高表达-低甲基化基因的热图;7B.14个低表达-高甲基化基因的热图注:“type”表示组织类型,“N”表示正常组织,“S”表示瘢痕组织2.4 统计分析结果
ROC曲线分析结果显示,CCR2、LMO7、STEAP4、NNAT、TCF7L2、ZC3H12B等基因具有较好的瘢痕诊断效能,其曲线下面积(AUC)值居于前列。见表2。
表2 烧伤后瘢痕组织与正常组织异常甲基化基因受试者操作特征曲线下面积及P值基因 曲线下面积 P值 CCR2 0.972 <0.001 LMO7 0.972 0.011 STEAP4 0.972 <0.001 NNAT 0.944 <0.001 TCF7L2 0.944 0.003 ZC3H12B 0.944 0.010 AMPD3 0.917 <0.001 BTBD17 0.917 0.039 PRICKLE1 0.917 <0.001 将这些基因按照AUC值从大到小依次纳入SVM模型中,当纳入CCR2、LMO7、STEAP4、NNAT和TCF7L2这5个基因时,惩罚参数c和核函数参数g都<0.01,此时构建的分类器模型分类准确率为93.3%,表明上述基因有较好的分类功能(图8)。
 8 烧伤后瘢痕组织差异表达甲基化基因的数据展示及5个差异甲基化基因可视图。8A、8B、8C、8D、8E、8F.分别为数据的整体展示及TCF7L2、NNAT、LMO7、CCR2和STEAP4基因的分维可视化图
8 烧伤后瘢痕组织差异表达甲基化基因的数据展示及5个差异甲基化基因可视图。8A、8B、8C、8D、8E、8F.分别为数据的整体展示及TCF7L2、NNAT、LMO7、CCR2和STEAP4基因的分维可视化图3. 讨论
烧伤是临床上常见的疾病,其中相当一部分患者为深Ⅱ度及以上烧伤。瘢痕是烧伤后康复期患者最常见的并发症之一,该疾病会严重影响患者的外貌和生活质量。因此,探究烧伤后瘢痕组织的形成机制并寻找潜在的治疗靶点对改善烧伤患者的预后具有重要意义。本研究利用WGCNA分析寻找与烧伤后瘢痕形成相关的基因模块,并探究了模块内的基因生物功能和甲基化状态,该研究为治疗烧伤后瘢痕提供了有意义的临床参考价值。
根据富集分析的结果,棕色模块内的基因具有“雄激素受体信号通路的调控”功能,因此本研究认为雄激素受体在瘢痕组织生成过程中具有一定作用;且由于DNA甲基化状态的改变,该通路状态发生改变。而Schierle等[9]研究结果显示,雄激素受体DNA的含量在瘢痕组织与正常组织之间具有显著差异;同时指出抗雄激素物质可通过竞争结合雄激素受体导致瘢痕组织中Ⅰ型前胶原mRNA低表达,因此本研究认为抗雄激素物质具有治疗瘢痕的潜力。“细胞因子-细胞因子受体相互作用”在KEGG分析中被富集到,研究表明多种细胞因子如血管源性生长因子、IL家族等对瘢痕生成有重要影响[10, 11, 12, 13]。因此本研究认为模块内的基因可调节细胞因子受体,与微环境内的细胞因子相互作用并诱导瘢痕组织的形成。Metascape数据库的分析显示,“胰岛素分泌的正调节”被显著富集,表明这些异常甲基化的基因可能导致胰岛素相关通路的改变。因此本研究认为胰岛素在瘢痕生成方面具有重要作用,Hallam等[14]进行的一项随机对照实验显示,皮下注射胰岛素可减少瘢痕产生。因此,本研究认为胰岛素注射也可作为瘢痕治疗的潜在疗法。
在棕色模块内共显示了35个异常甲基化基因,其中CCR2、LMO7、STEAP4、NNAT和TCF7L2具有良好的瘢痕诊断效能并被纳入到SVM模型中。CCR2编码单核细胞趋化蛋白-1,Frik等[15]研究显示CCR2敲除小鼠脑损伤后单核细胞浸润减少且瘢痕形成减少。本研究中观察到瘢痕组织中CCR2是高表达的,推测CCR2可诱导瘢痕产生,是瘢痕治疗的潜在靶点。本研究结果显示LMO7在瘢痕组织中高表达,推测LMO7可促进瘢痕组织产生。Xie等[16]研究表明LMO7由TGF-β诱导并通过TGF-β途径负反馈调节在伤口愈合和瘢痕生成中发挥重要作用,这与本研究结果一致。STEAP4、NNAT和TCF7L2则未见相应报道,其在瘢痕产生中的作用可能需要进一步实验研究进行验证。
总之,本研究通过WGCNA初步分析了烧伤后瘢痕产生的分子机制和异常甲基化基因,结果显示基因CCR2、LMO7、STEAP4、NNAT和TCF7L2在瘢痕形成中具有重要作用,可作为治疗的潜在靶点。
何佳、王婧薷:酝酿和设计实验、实验研究和论文撰写;甘文军、李桂强:数据整理、分析数据;信琪、林泽鹏、阮树斌:统计分析;陈晓东:研究指导、经费支持所有作者均声明不存在利益冲突 -
参考文献
(22) [1] RodriguesM,KosaricN,BonhamCA,et al.Wound healing: a cellular perspective[J].Physiol Rev,2019,99(1):665-706.DOI: 10.1152/physrev.00067.2017. [2] StegleO,TeichmannSA,MarioniJC.Computational and analytical challenges in single-cell transcriptomics[J].Nat Rev Genet,2015,16(3):133-145.DOI: 10.1038/nrg3833. [3] PatelS,SrivastavaS,SinghMR,et al.Mechanistic insight into diabetic wounds: pathogenesis, molecular targets and treatment strategies to pace wound healing[J].Biomed Pharmacother,2019,112:108615.DOI: 10.1016/j.biopha.2019.108615. [4] NielsenJS,McNagnyKM.Novel functions of the CD34 family[J].J Cell Sci,2008,121(Pt 22):3683-3692.DOI: 10.1242/jcs.037507. [5] Díaz-FloresL,GutiérrezR,GarcíaMP,et al.Cd34 + stromal cells/telocytes in normal and pathological skin[J].Int J Mol Sci,2021,22(14):7342.DOI: 10.3390/ijms22147342. [6] WangL,QinW,ZhouY,et al.Transforming growth factor β plays an important role in enhancing wound healing by topical application of Povidone-iodine[J].Sci Rep,2017,7(1):991.DOI: 10.1038/s41598-017-01116-5. [7] ZhuB,NahmiasY,YarmushML,et al.Microfluidic isolation of CD34-positive skin cells enables regeneration of hair and sebaceous glands in vivo[J].Stem Cells Transl Med,2014,3(11):1354-1362.DOI: 10.5966/sctm.2014-0098. [8] NuutilaK,SinghM,KruseC,et al.Wound healing from dermal grafts containing CD34 + cells is comparable to wound healing with split-thickness skin micrografts[J].Plast Reconstr Surg,2017,140(2):306-314.DOI: 10.1097/PRS.0000000000003516. [9] HeJ,HuangW,WangJ,et al.Single-cell analysis reveals distinct functional heterogeneity of CD34 + cells in anagen wound and diabetic wound[J].Biochem Biophys Res Commun,2023,639:9-19.DOI: 10.1016/j.bbrc.2022.11.080. [10] HanJ,LinK,ChooH,et al.β-catenin signaling evokes hair follicle senescence by accelerating the differentiation of hair follicle mesenchymal progenitors[J].Front Cell Dev Biol,2022,10:839519.DOI: 10.3389/fcell.2022.839519. [11] PennigJ,ScherrerP,GisslerMC,et al.Glucose lowering by SGLT2-inhibitor empagliflozin accelerates atherosclerosis regression in hyperglycemic STZ-diabetic mice[J].Sci Rep,2019,9(1):17937.DOI: 10.1038/s41598-019-54224-9. [12] LandénNX,LiD,StåhleM.Transition from inflammation to proliferation: a critical step during wound healing[J].Cell Mol Life Sci,2016,73(20):3861-3885.DOI: 10.1007/s00018-016-2268-0. [13] SunZ,SunX,YuanY,et al.FCGR2B as a prognostic and immune microenvironmental marker for gliomas based on transcriptomic analysis[J].Medicine (Baltimore),2023,102(37):e35084.DOI: 10.1097/MD.0000000000035084. [14] DevosH,ZoidakisJ,RoubelakisMG,et al.Reviewing the regulators of COL1A1[J].Int J Mol Sci,2023,24(12):10004.DOI: 10.3390/ijms241210004. [15] TaiY,WoodsEL,DallyJ,et al.Myofibroblasts: function, formation, and scope of molecular therapies for skin fibrosis[J].Biomolecules,2021,11(8):1095.DOI: 10.3390/biom11081095. [16] GanQ,YoshidaT,LiJ,et al.Smooth muscle cells and myofibroblasts use distinct transcriptional mechanisms for smooth muscle alpha-actin expression[J].Circ Res,2007,101(9):883-892.DOI: 10.1161/CIRCRESAHA.107.154831. [17] DanielJM,ProckA,DutzmannJ,et al.Regulator of G-protein signaling 5 prevents smooth muscle cell proliferation and attenuates neointima formation[J].Arterioscler Thromb Vasc Biol,2016,36(2):317-327.DOI: 10.1161/ATVBAHA.115.305974. [18] ChenZ,ZhouL,LiuL,et al.Single-cell RNA sequencing highlights the role of inflammatory cancer-associated fibroblasts in bladder urothelial carcinoma[J].Nat Commun,2020,11(1):5077.DOI: 10.1038/s41467-020-18916-5. [19] SreedharA,Aguilera-AguirreL,SinghKK.Mitochondria in skin health, aging, and disease[J].Cell Death Dis,2020,11(6):444.DOI: 10.1038/s41419-020-2649-z. [20] AkermanSC,HossainS,ShoboA,et al.Neurodegenerative disease-related proteins within the epidermal layer of the human skin[J].J Alzheimers Dis,2019,69(2):463-478.DOI: 10.3233/JAD-181191. [21] NodaS,HosoyaT,KomiyaY,et al.CD34 +THY1 + synovial fibroblast subset in arthritic joints has high osteoblastic and chondrogenic potentials in vitro[J].Arthritis Res Ther,2022,24(1):45.DOI: 10.1186/s13075-022-02736-7. [22] DuZ,YouX,WuD,et al.Rhythm disturbance in osteoarthritis[J].Cell Commun Signal,2022,20(1):70.DOI: 10.1186/s12964-022-00891-7. -
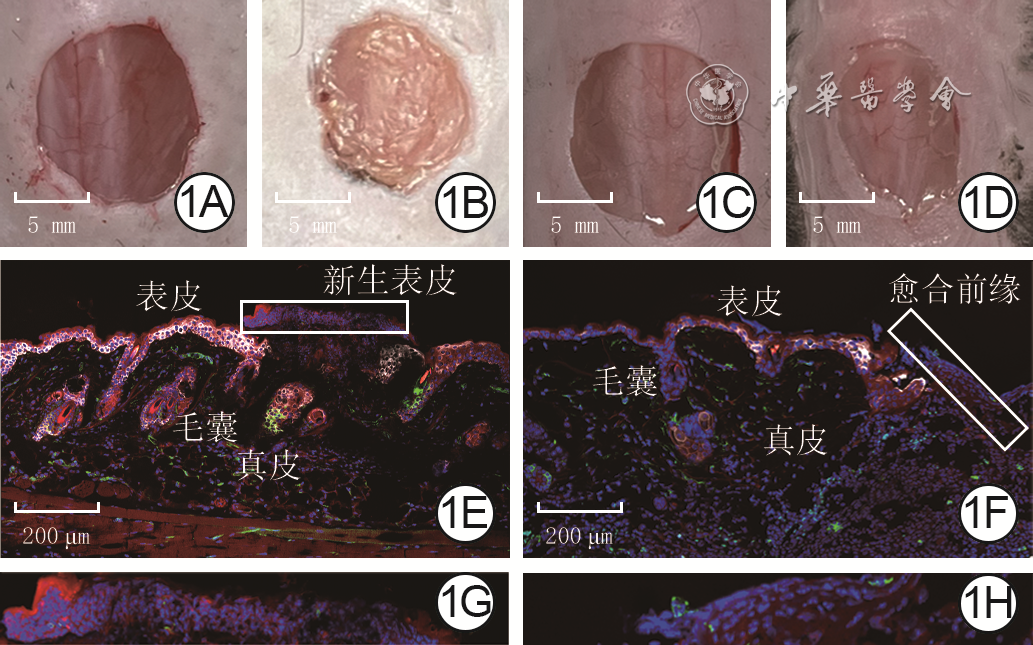
1 2组全层皮肤缺损小鼠伤后各时间点创面形态及伤后4 d CD34 +细胞在创面中的定位。1A、1B.分别为对照组伤后0(即刻)、4 d创面形态,图1B中创面面积缩小,表皮再上皮化;1C、1D.分别为糖尿病组伤后0、4 d创面形态,图1D创面较图1B愈合延迟,仅有较薄的新生表皮;1E、1F.分别为CD34 +细胞在对照组、糖尿病组创面中的定位以及与角蛋白14共定位,CD34 +细胞位于创面周围组织的真皮、毛囊、血管周围和部分表皮细胞以及创面新生组织,在图1F创面周围组织的毛囊中减少 Alexa Fluor 647-4',6-二脒基-2-苯基吲哚×20;1G、1H.分别为图1E、1F中方框中图形的放大图,可见CD34表达在创面新生表皮的部分细胞中 Alexa Fluor 647-4',6-二脒基-2-苯基吲哚×60
注:对照组小鼠为CD34+细胞谱系追踪小鼠,糖尿病组小鼠为CD34+细胞谱系追踪糖尿病小鼠;CD34+细胞呈绿色,非CD34+细胞呈红色,角蛋白14呈白色,细胞核呈蓝色
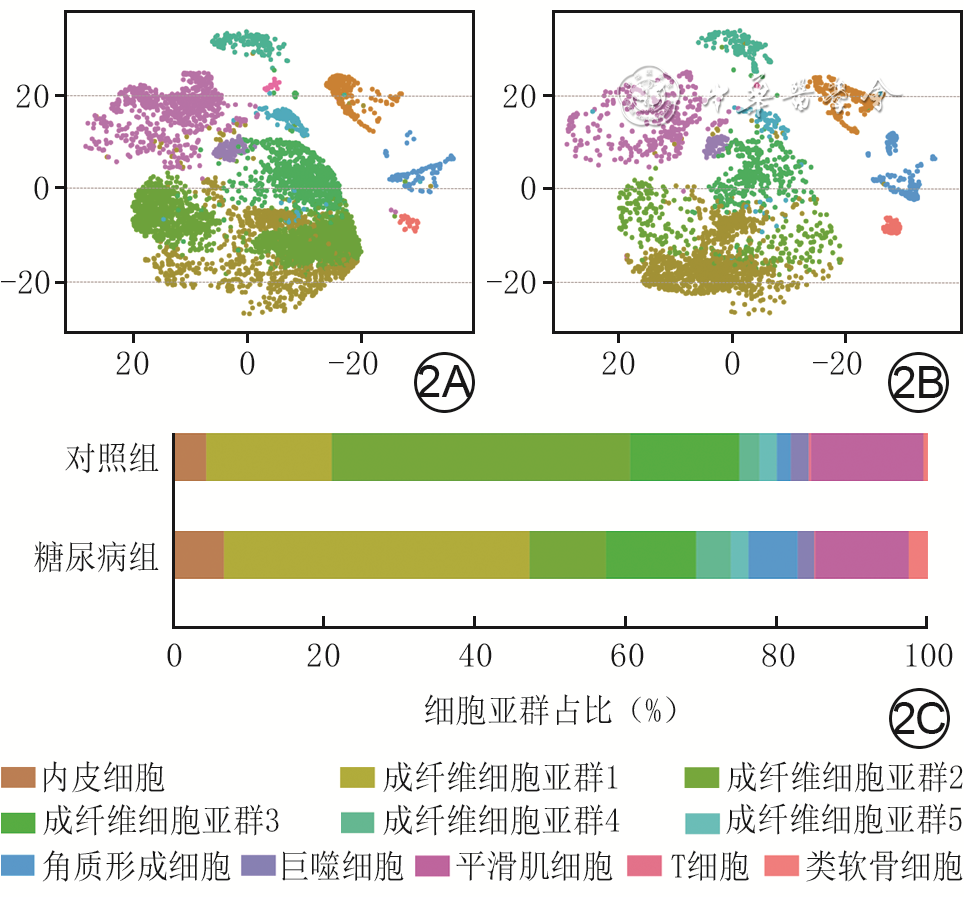
2 2组全层皮肤缺损小鼠伤后4 d创面组织中CD34 +细胞亚群及其占比。2A、2B.分别为对照组、糖尿病组创面中CD34 +细胞亚群的二维流形近似和投影云图;2C.CD34 +细胞亚群占比
注:对照组小鼠为CD34+细胞谱系追踪小鼠,糖尿病组小鼠为CD34+细胞谱系追踪糖尿病小鼠
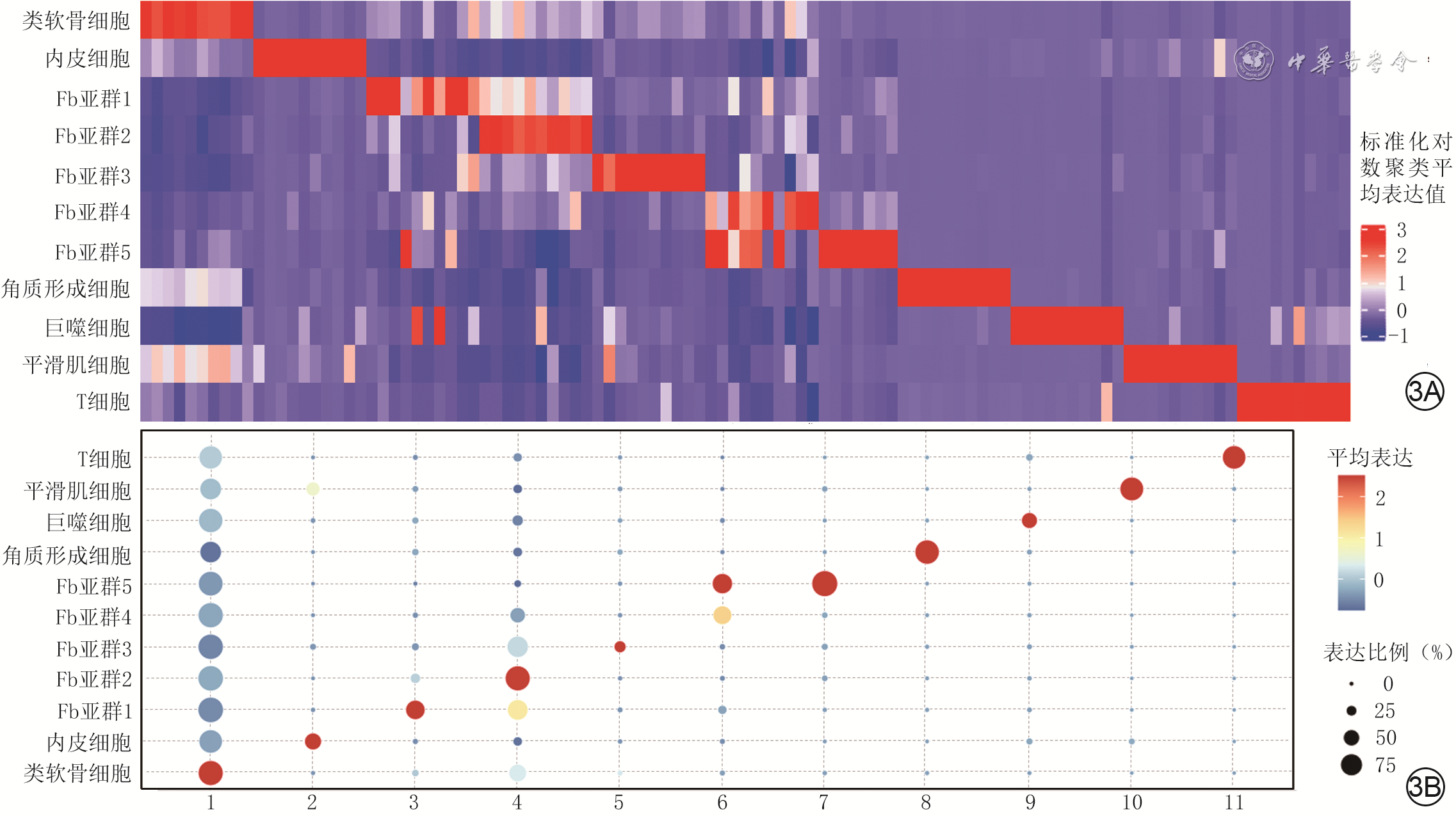
3 总体分析2组全层皮肤缺损小鼠伤后4 d创面组织中各CD34 +细胞亚群前10高表达标记基因及注释细胞亚群的标记基因。3A.热图展示各CD34 +细胞亚群前10高表达标记基因;3B.气泡图展示注释细胞亚群的标记基因
注:对照组小鼠为CD34+细胞谱系追踪小鼠,糖尿病组小鼠为CD34+细胞谱系追踪糖尿病小鼠;图3A从左至右各列依次分别为标记类软骨细胞的线粒体编码的细胞色素C氧化酶Ⅱ(MT-CO2)、MT-CO1、MT-CO3、线粒体细胞色素b、线粒体编码的烟酰胺腺嘌呤二核苷酸:泛醌氧化还原酶核心亚基1(MT-ND1)、线粒体编码的ATP合酶膜亚基6、MT-ND4、MT-ND2、MT-ND3、转移相关肺腺癌转录本1(MALAT1),标记内皮细胞的脂肪酸结合蛋白4(FABP4)、选择素P、血小板和内皮细胞黏附分子1、水通道蛋白1、CD36分子、跨膜4 L六家族成员1、质膜小囊泡相关蛋白、清道夫受体B类成员1、含载脂蛋白L结构域1、富含亮氨酸的α-2-糖蛋白1,标记成纤维细胞(Fb)亚群1的Gremlin 1(GREM1)、胰岛素样生长因子结合蛋白1、基质金属蛋白酶3(MMP3)、细胞视黄酸结合蛋白1、Fcγ受体Ⅱb、MMP11、凝血因子ⅩⅢ A链、ⅩⅩⅢ型胶原蛋白α1(COL23A1)、羧肽酶X-M14家族成员1、血清淀粉样蛋白A3,标记Fb亚群2的补体成分4B(C4B)、羧肽酶Z、COL1A2、COL1A1、AE结合蛋白1、转化生长因子β诱导蛋白、类纤维蛋白原样蛋白2、前胶原C-内肽酶增强子2、整合素亚基β样1、导向蛋白3B抗体,标记Fb亚群3的H19印记母源表达转录本(H19)、C-X-C趋化因子配体1(CXCL1)、CXCL5、正五聚蛋白3、分泌型卷曲相关蛋白2、微纤维相关蛋白5、白细胞介素(IL)1受体类似物2、蛋白聚糖4、血小板反应蛋白4和COL4A1,标记Fb亚群4的Dickkopf Wnt信号通路抑制剂2(DKK2)、外核苷酸焦磷酸酶/磷酸二酯酶2、凝血因子Ⅱ凝血酶受体、肽酶抑制剂16、细胞色素P450家族1亚家族B成员1、生长停滞特异性1、群体凝血因子C同源物、核受体亚家族4A组成员1、弹性蛋白、结缔组织生长因子,标记Fb亚群5的纤维调节蛋白(FMOD)、Wnt抑制因子1、腱调蛋白、胆囊收缩素、胶质蛋白、Ig超家族成员10(IGSF10)和IGSF5,标记角质形成细胞的角蛋白5、分层蛋白、角蛋白17、角蛋白15、角蛋白14、半乳糖凝集素7、与外周髓磷脂蛋白22相关的P53凋亡效应物质、胎盘表达转录本1、角蛋白6a和角质蛋白连接蛋白,标记巨噬细胞的CD74分子、组织相容性Ⅱ类抗原Aα、H2-Ab1、溶菌酶2、C-C型趋化因子配体3(CCL3)、S100钙结合蛋白A9、CCL4、CD14分子、丝氨酸和CXCL3,标记平滑肌细胞的G蛋白信号调节蛋白5(RGS5)、Notch受体3、肌球蛋白重链11、11号染色体开放阅读框96、内皮素受体B型、肌球蛋白轻链9、平滑肌肌动蛋白α-2、肌球蛋白轻链激酶、富含半胱氨酸的酸性分泌蛋白类似蛋白1、转胶蛋白,标记T细胞的可诱导T细胞共刺激分子(ICOS)、IL-17A、IL-17F、RGS1、IL-2受体亚单位β、溶酶体蛋白跨膜5、胚细胞抗原、环腺苷酸反应元件调制器、液泡蛋白分拣相关蛋白37B、RGS2;图3B横坐标下1为MALAT1,2为FABP4,3为GREM1,4为C4B,5为H19,6为DKK2,7为FMOD,8为角蛋白5,9为CD74分子,10为RGS5,11为ICOS;标记Fb亚群5的3个基因与其他细胞标记基因重叠
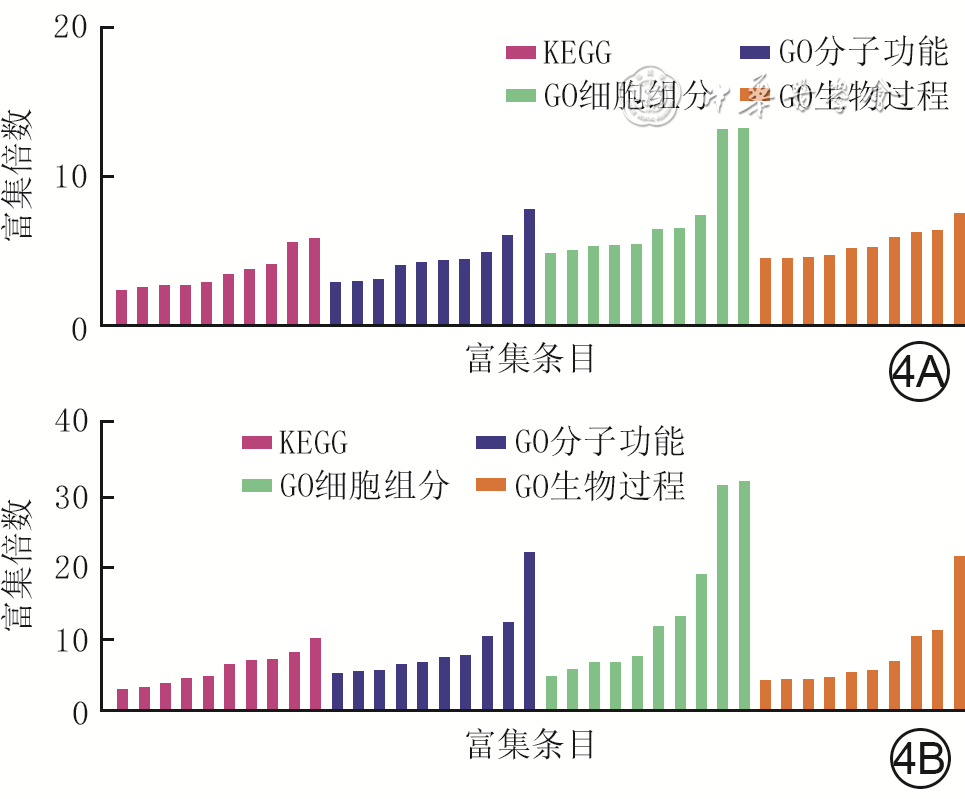
4 2组全层皮肤缺损小鼠伤后4 d创面组织间CD34 +成纤维细胞亚群1差异表达显著的DEG的GO和KEGG富集分析。4A.糖尿病组较对照组显著上调的DEG;4B.糖尿病组较对照组显著下调的DEG
注:对照组小鼠为CD34+细胞谱系追踪小鼠,糖尿病组小鼠为CD34+细胞谱系追踪糖尿病小鼠;KEGG为京都基因与基因组百科全书,GO为基因本体论,DEG为差异表达基因;图4A横坐标从左至右对应富集条目为破骨细胞分化、白细胞介素17(IL-17)信号通路、丝裂原活化蛋白激酶信号通路、抗原处理和呈递、Janus激酶-信号转导及转录活化因子信号通路、肿瘤坏死因子信号通路、细胞因子-细胞因子受体相互作用、核糖体、流体剪切应力与动脉粥样硬化、冠状病毒疾病-新型冠状病毒感染、蛋白质同源二聚活性、肽酶活性、金属内肽酶活性、相同蛋白质结合、蛋白酶结合、金属肽酶活性、肽酶抑制剂活性、核糖体的结构成分、受体结合、蛋白质结合、活化蛋白-1转录复合物、细胞外基质、核糖体、细胞表面、细胞核、核糖核蛋白复合体、细胞质、胞质核糖体、细胞外区、细胞外间隙、正调控血管生成、动物器官再生、肽酶活性的负调控、蛋白质加工、细胞对IL-1的反应、对肽酶激素的反应、细胞迁移的正向调节、对脂多糖的反应、细胞增殖的负调控、细胞翻译;图4B横坐标从左至右对应富集条目为阿米巴病、库欣综合征、EB病毒感染、磷脂酰肌醇3-激酶-蛋白激酶B信号通路、糖尿病并发症中的晚期糖基化终末产物-晚期糖基化终末产物受体信号通路、黏着斑、人乳头瘤病毒感染、癌症中的蛋白聚糖、细胞外基质受体相互作用、蛋白质的消化和吸收、肝素结合、蛋白酶结合、相同蛋白质结合、血小板衍生生长因子结合、整合素结合、钙离子结合、蛋白质结合、胶原结合、赋予拉伸强度的细胞外基质结构成分、细胞外基质结构成分、弹性纤维、内质网、细胞表面、肌膜、肌腱、基底膜、胶原三聚体、细胞外间隙、细胞外区、细胞外基质、炎症反应的负调控、成纤维细胞生长因子受体信号通路的负调控、细胞迁移、细胞对干扰素β的反应、胶原生物合成过程、细胞-基质黏附、创面修复、细胞黏附、胶原原纤维组装、细胞外基质组装
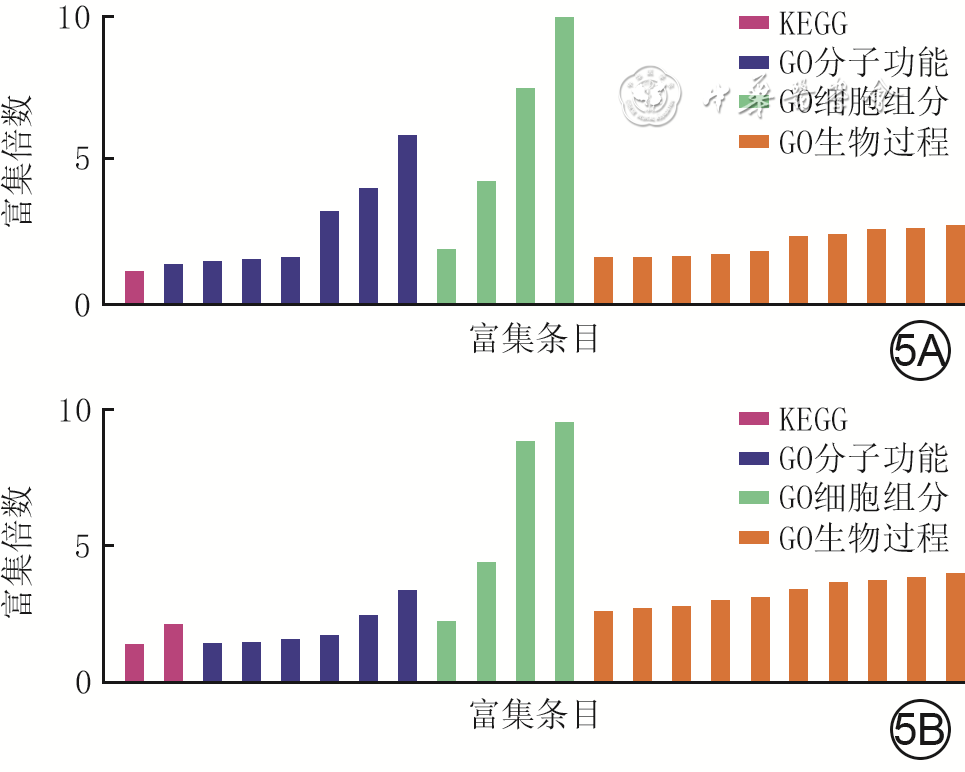
5 2组全层皮肤缺损小鼠伤后4 d创面组织间CD34 +成纤维细胞亚群2差异表达显著的DEG的GO和KEGG富集分析。5A.糖尿病组较对照组显著上调的DEG;5B.糖尿病组较对照组显著下调的DEG
注:对照组小鼠为CD34+细胞谱系追踪小鼠,糖尿病组小鼠为CD34+细胞谱系追踪糖尿病小鼠;KEGG为京都基因与基因组百科全书,GO为基因本体论,DEG为差异表达基因;图5A横坐标从左至右对应富集条目为癌症中的微小RNA、白细胞介素17信号通路、序列特异性双链DNA结合、肽酶活性、鸟苷三磷酸酶活性、金属肽酶活性、双链DNA结合、肝素结合、高密度脂蛋白颗粒、细胞外基质(ECM)、细胞外间隙、细胞外区、衰老、细胞对过氧化氢的反应、短期记忆、胶原原纤维组装、细胞增殖调控、细胞凋亡过程的正调控、正调控血管生成、对有机环状化合物的反应、对氧化应激的反应、细胞增殖的负调控;图5B横坐标从左至右对应富集条目为代谢途径、糖胺聚糖结合、肝素结合、钙离子结合、生长因子活性、胰岛素样生长因子结合、生长因子结合、ECM结构成分、基底膜、ECM、细胞外间隙、细胞外区、内皮细胞形态发生、细胞增殖的负调控、对维生素D的反应、钙离子转运的负调控、转化生长因子β产生的正调控、对糖皮质激素的反应、细胞对环腺苷酸的反应、细胞增殖的正向调节、ECM组装、衰老
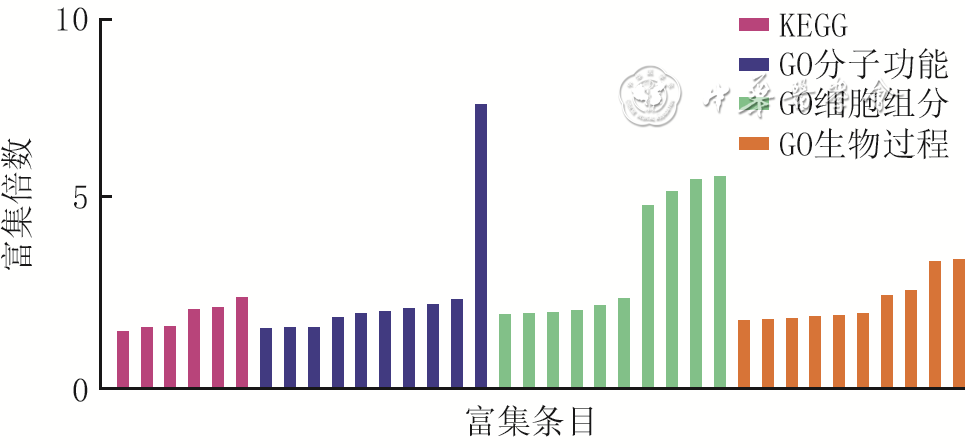
6 较对照组显著上调的糖尿病组小鼠伤后4 d全层皮肤缺损创面组织中CD34 +平滑肌细胞差异表达基因的GO和KEGG富集分析
注:对照组小鼠为CD34+细胞谱系追踪小鼠,糖尿病组小鼠为CD34+细胞谱系追踪糖尿病小鼠;KEGG为京都基因与基因组百科全书,GO为基因本体论;横坐标从左至右对应富集条目为内吞作用、流体剪切应力与动脉粥样硬化、人巨噬细胞感染、癌症中的蛋白聚糖、癌症中的发病途径、吞噬体、整合素结合、蛋白激素结合、肽酶活性、相同蛋白质结合、泛素-蛋白质连接酶结合、金属离子结合、前mRNA内含子结合、酶结合、细胞外基质结合、蛋白质结合、线粒体、RNA聚合酶Ⅱ转录因子复合物、核基质、高尔基膜、核糖核蛋白复合体、内质网、核、细胞质、核质、细胞溶质、神经元凋亡过程的正调控、神经元投射发育的负调控、细胞质微管组织、镁离子传输、细胞外基质组装、信号肽处理、转录的正调控(DNA模板)、骨骼系统发育、凋亡过程、细胞迁移
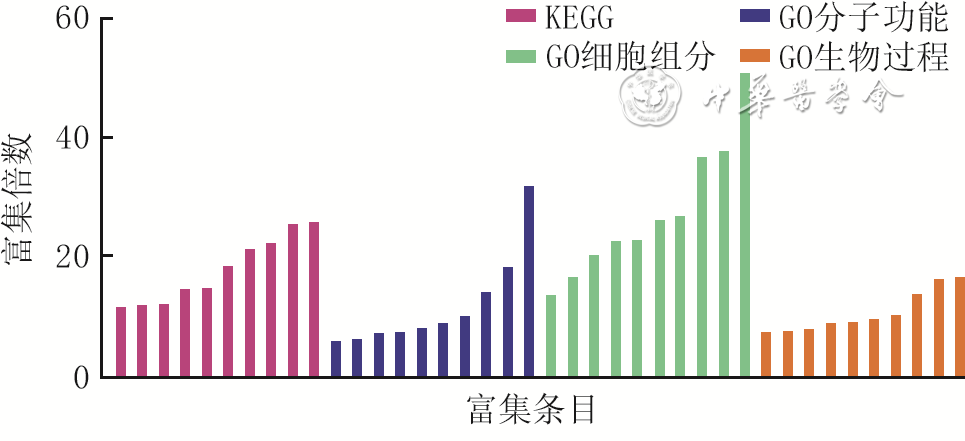
7 2组全层皮肤缺损小鼠伤后4 d创面组织间CD34 +角质形成细胞差异表达显著的差异表达基因的GO和KEGG富集分析
注:对照组小鼠为CD34+细胞谱系追踪小鼠,糖尿病组小鼠为CD34+细胞谱系追踪糖尿病小鼠;KEGG为京都基因与基因组百科全书,GO为基因本体论;横坐标从左至右对应富集条目为非酒精性脂肪肝、糖尿病性心肌病、化学致癌-活性氧、氧化磷酸化、神经退行性变的途径、脊髓侧索硬化症、亨廷顿舞蹈症、阿尔兹海默病、帕金森病、朊病毒病、烟酰胺腺嘌呤二核苷酸(泛醌)活性、核苷酸活性、核酸结合、相同蛋白质结合、单链DNA结合、大分子复合物结合、酶结合、染色质结合、RNA结合、蛋白质结合、剪切体复合物、线粒体呼吸链复合体Ⅰ、呼吸链、核糖核蛋白复合体、细胞溶质、线粒体内膜、线粒体、细胞质、核质、细胞核、线粒体电子运输、DNA复制、核糖体RNA处理、信使RNA剪切(通过剪切体)、细胞周期、转录的正调控(DNA模板)、线粒体呼吸链复合体Ⅰ组装、有氧呼吸、RNA剪切、线粒体ATP合成偶联质子转运
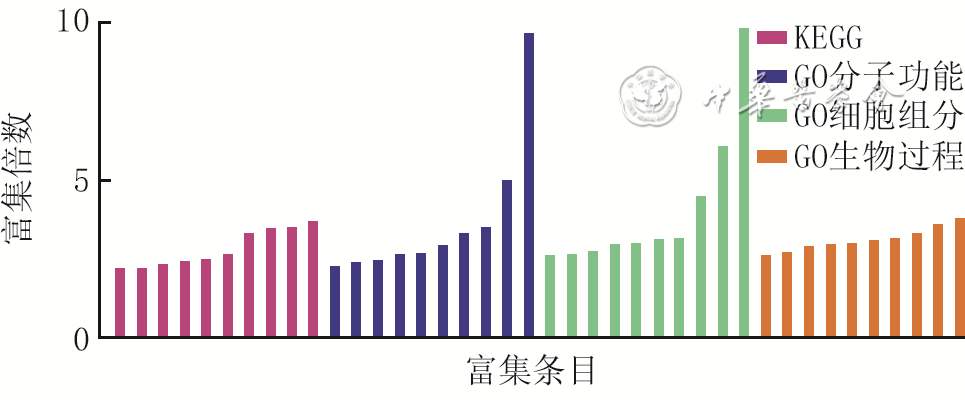
8 2组全层皮肤缺损小鼠伤后4 d创面组织间CD34 +类软骨细胞差异表达显著的差异表达基因的GO和KEGG富集分析
注:对照组小鼠为CD34+细胞谱系追踪小鼠,糖尿病组小鼠为CD34+细胞谱系追踪糖尿病小鼠;KEGG为京都基因与基因组百科全书,GO为基因本体论;横坐标从左至右对应富集条目为阿米巴病、化学致癌-活性氧、糖尿病并发症中晚期糖基化终末产物(AGE)-AGE受体信号、内吞作用、流体剪切应力与动脉粥样硬化、抗原处理和呈递、糖尿病性心肌病、癌症中的蛋白聚糖、爱泼斯坦-巴尔病毒感染、卡波西肉瘤相关疱疹病毒感染、染色体DNA结合、整合素结合、细胞外基质(ECM)结构成分、肽酶活性、相同蛋白质结合、酶结合、泛素蛋白连接酶结合、胶原结合、大分子复合物结合、蛋白质结合、核内小体、大分子化合物、核糖核蛋白复合体、细胞表面、细胞外区、早期内膜体、ECM、核质、细胞质、细胞核、自然杀伤细胞介导的细胞毒性的保护、对过氧化氢的反应、蛋白质磷酸化的正调控、细胞迁移的正向调控、基因表达的昼夜节律调节、染色质组装、昼夜节律的调控、内皮细胞迁移的正调控、转录的负调控(DNA模板)、节律过程
-

 下载:
下载:

 下载:
下载:

计量
- 文章访问数: 8027
- HTML全文浏览量: 67
- PDF下载量: 54
- 被引次数: 0
