- Medline/PubMed数据库
- Scopus数据库
- PMC数据库
- CSCD
- 北大核心收录期刊
- 统计源期刊
- 我国高质量科技期刊T1级
- 第6届中国精品科技期刊
| Citation: | Khoong YM,Huang X,Meng XC,et al.Effects of transient receptor potential vanilloid type 4-specific activator on human vascular endothelial cell functions and blood supply of rat perforator flap and its mechanism[J].Chin J Burns Wounds,2022,38(5):434-446.DOI: 10.3760/cma.j.cn501120-20210419-00138.

|

| [1] |
SmithRK,WykesJ,MartinDT,et al.Perforator variability in the anterolateral thigh free flap: a systematic review[J].Surg Radiol Anat,2017,39(7):779-789.DOI: 10.1007/s00276-016-1802-y.
|
| [2] |
LieKH,BarkerAS,AshtonMW.A classification system for partial and complete DIEP flap necrosis based on a review of 17,096 DIEP flaps in 693 articles including analysis of 152 total flap failures[J].Plast Reconstr Surg,2013,132(6):1401-1408.DOI: 10.1097/01.prs.0000434402.06564.bd.
|
| [3] |
RiniIS,GunardiAJ,MarsaulinaRP,et al.A systematic review of the keystone design perforator island flap in the reconstruction of trunk defects[J].Arch Plast Surg,2020,47(6):535-541.DOI: 10.5999/aps.2020.00094.
|
| [4] |
JiangZM, LiXC, ChenM, et al. Effect of endogenous vascular endothelial growth factor on flap surgical delay in a rat flap model[J]. Plast Reconstr Surg, 2019, 143(1): 126-135. DOI: 10.1097/PRS.0000000000005145.
|
| [5] |
TaylorGI, CorlettRJ, CaddyCM,et al. An anatomic review of the delay phenomenon: Ⅱ. Clinical applications [J]. Plast Reconstr Surg, 1992, 89(3): 408-416;discussion 417-418.
|
| [6] |
MiyamotoS,MinabeT,HariiK.Effect of recipient arterial blood inflow on free flap survival area[J].Plast Reconstr Surg,2008,121(2):505-513.DOI: 10.1097/01.prs.0000299185.32881.55.
|
| [7] |
TaylorGI,ChubbDP,AshtonMW.True and 'choke' anastomoses between perforator angiosomes: part Ⅰ. Anatomical location[J].Plast Reconstr Surg,2013,132(6):1447-1456.DOI: 10.1097/PRS.0b013e3182a80638.
|
| [8] |
DharSC,TaylorGI.The delay phenomenon: the story unfolds[J].Plast Reconstr Surg,1999,104(7):2079-2091.DOI: 10.1097/00006534-199912000-00021.
|
| [9] |
GhaliS, ButlerPEM, TepperOM, et al. Vascular delay revisited[J]. Plast Reconstr Surg, 2007, 119(6): 1735-1744. DOI: 10.1097/01.prs.0000246384.14593.6e.
|
| [10] |
LuoZH,WuPF,QingLM,et al.The hemodynamic and molecular mechanism study on the choke vessels in the multi-territory perforator flap transforming into true anastomosis[J].Gene,2019,687:99-108.DOI: 10.1016/j.gene.2018.11.019.
|
| [11] |
QingLM,LeiPF,TangJY,et al.Inflammatory response associated with choke vessel remodeling in the extended perforator flap model[J].Exp Ther Med,2017,13(5):2012-2018.DOI: 10.3892/etm.2017.4205.
|
| [12] |
GaliePA,NguyenDH,ChoiCK,et al.Fluid shear stress threshold regulates angiogenic sprouting[J].Proc Natl Acad Sci U S A,2014,111(22):7968-7973.DOI: 10.1073/pnas.1310842111.
|
| [13] |
SeilerC,StollerM,PittB,et al.The human coronary collateral circulation: development and clinical importance[J].Eur Heart J,2013,34(34):2674-2682.DOI: 10.1093/eurheartj/eht195.
|
| [14] |
HendrikseJ,HartkampMJ,HillenB,et al.Collateral ability of the circle of Willis in patients with unilateral internal carotid artery occlusion: border zone infarcts and clinical symptoms[J].Stroke,2001,32(12):2768-2773.DOI: 10.1161/hs1201.099892.
|
| [15] |
HoeferIE, van RoyenN,BuschmannIR,et al.Time course of arteriogenesis following femoral artery occlusion in the rabbit[J].Cardiovasc Res,2001,49(3):609-617.DOI: 10.1016/s0008-6363(00)00243-1.
|
| [16] |
SchierlingW,TroidlK,ApfelbeckH,et al.Cerebral arteriogenesis is enhanced by pharmacological as well as fluid-shear-stress activation of the Trpv4 calcium channel[J].Eur J Vasc Endovasc Surg,2011,41(5):589-596.DOI: 10.1016/j.ejvs.2010.11.034.
|
| [17] |
TroidlC,TroidlK,SchierlingW,et al.Trpv4 induces collateral vessel growth during regeneration of the arterial circulation[J].J Cell Mol Med,2009,13(8B):2613-2621.DOI: 10.1111/j.1582-4934.2008.00579.x.
|
| [18] |
BubolzAH,MendozaSA,ZhengXD,et al.Activation of endothelial TRPV4 channels mediates flow-induced dilation in human coronary arterioles: role of Ca2+ entry and mitochondrial ROS signaling[J].Am J Physiol Heart Circ Physiol,2012,302(3):H634-642.DOI: 10.1152/ajpheart.00717.2011.
|
| [19] |
SonkusareSK,BonevAD,LedouxJ,et al.Elementary Ca2+ signals through endothelial TRPV4 channels regulate vascular function[J].Science,2012,336(6081):597-601.DOI: 10.1126/science.1216283.
|
| [20] |
DarbyWG,PotocnikS,RamachandranR,et al.Shear stress sensitizes TRPV4 in endothelium-dependent vasodilatation[J].Pharmacol Res,2018,133:152-159.DOI: 10.1016/j.phrs.2018.05.009.
|
| [21] |
BaeJH,WangZC,LiHZ,et al.Activation of TRPV4 increases neovascularization of rat prefabricated flaps[J].J Reconstr Microsurg,2018,34(1):35-40.DOI: 10.1055/s-0037-1607210.
|
| [22] |
Welch-ReardonKM,WuN,HughesCCW.A role for partial endothelial-mesenchymal transitions in angiogenesis?[J].Arterioscler Thromb Vasc Biol,2015,35(2):303-308.DOI: 10.1161/ATVBAHA.114.303220.
|
| [23] |
HultgrenNW,FangJS,ZieglerME,et al.Slug regulates the Dll4-Notch-VEGFR2 axis to control endothelial cell activation and angiogenesis[J].Nat Commun,2020,11(1):5400.DOI: 10.1038/s41467-020-18633-z.
|
| [24] |
AzimiI,RobitailleM,ArmitageK,et al.Activation of the ion channel TRPV4 induces epithelial to mesenchymal transition in breast cancer cells[J].Int J Mol Sci,2020,21(24):9417.DOI: 10.3390/ijms21249417.
|
| [25] |
CappelliHC,KanugulaAK,AdapalaRK,et al.Mechanosensitive TRPV4 channels stabilize VE-cadherin junctions to regulate tumor vascular integrity and metastasis[J].Cancer Lett,2019,442:15-20.DOI: 10.1016/j.canlet.2018.07.042.
|
| [26] |
WangHF, ZhangBY, WangX, et al. TRPV4 overexpression promotes metastasis through epithelial-mesenchymal transition in gastric cancer and correlates with poor prognosis[J]. Onco Targets Ther, 2020, 13: 8383-8394. DOI: 10.2147/OTT.S256918.
|
| [27] |
LeeWH,ChoongLY,JinTH,et al.TRPV4 plays a role in breast cancer cell migration via Ca2+-dependent activation of AKT and downregulation of E-cadherin cell cortex protein[J].Oncogenesis,2017,6(5):e338.DOI: 10.1038/oncsis.2017.39.
|
| [28] |
SharmaS,GoswamiR,RahamanSO.The TRPV4-TAZ mechanotransduction signaling axis in matrix stiffness- and TGFβ1-induced epithelial-mesenchymal transition[J].Cell Mol Bioeng,2019,12(2):139-152.DOI: 10.1007/s12195-018-00565-w.
|
| [29] |
SharmaS,GoswamiR,ZhangDX,et al.TRPV4 regulates matrix stiffness and TGFβ1-induced epithelial-mesenchymal transition[J].J Cell Mol Med,2019,23(2):761-774.DOI: 10.1111/jcmm.13972.
|
| [30] |
ZanT,LiHZ,HuangX,et al.Augmentation of perforator flap blood supply with sole or combined vascular supercharge and flap prefabrication for difficult head and neck reconstruction[J].Facial Plast Surg Aesthet Med,2020,22(6):441-448.DOI: 10.1089/fpsam.2020.0040.
|
| [31] |
LeseI,GrafDA,TsaiC,et al.Bioactive nanoparticle-based formulations increase survival area of perforator flaps in a rat model[J].PLoS One,2018,13(11):e0207802.DOI: 10.1371/journal.pone.0207802.
|
| [32] |
ChenM,LiXC,JiangZM,et al.Visualizing the pharmacologic preconditioning effect of botulinum toxin type a by infrared thermography in a rat pedicled perforator island flap model[J].Plast Reconstr Surg,2019,144(6):1016e-1024e.DOI: 10.1097/PRS.0000000000006251.
|
| [33] |
SilvestreJS,SmadjaDM,LévyBI.Postischemic revascularization: from cellular and molecular mechanisms to clinical applications[J].Physiol Rev,2013,93(4):1743-1802.DOI: 10.1152/physrev.00006.2013.
|
| [34] |
OlejarzW,Kubiak-TomaszewskaG,ChrzanowskaA,et al.Exosomes in angiogenesis and anti-angiogenic therapy in cancers[J].Int J Mol Sci,2020,21(16):5840.DOI: 10.3390/ijms21165840.
|
| [35] |
CarmelietP.Mechanisms of angiogenesis and arteriogenesis[J].Nat Med,2000,6(4):389-395.DOI: 10.1038/74651.
|
| [36] |
AdapalaRK,ThoppilRJ,GhoshK,et al.Activation of mechanosensitive ion channel TRPV4 normalizes tumor vasculature and improves cancer therapy[J].Oncogene,2016,35(3):314-322.DOI: 10.1038/onc.2015.83.
|
| [37] |
ThoppilRJ,CappelliHC,AdapalaRK,et al.TRPV4 channels regulate tumor angiogenesis via modulation of Rho/Rho kinase pathway[J].Oncotarget,2016,7(18):25849-25861.DOI: 10.18632/oncotarget.8405.
|
| [38] |
WenL,WenYC,KeGJ,et al.TRPV4 regulates migration and tube formation of human retinal capillary endothelial cells[J].BMC Ophthalmol,2018,18(1):38.DOI: 10.1186/s12886-018-0697-2.
|
| [39] |
TilletE,VittetD,FéraudO,et al.N-cadherin deficiency impairs pericyte recruitment, and not endothelial differentiation or sprouting, in embryonic stem cell-derived angiogenesis[J].Exp Cell Res,2005,310(2):392-400.DOI: 10.1016/j.yexcr.2005.08.021.
|
| [40] |
Welch-ReardonKM,EhsanSM,WangK,et al.Angiogenic sprouting is regulated by endothelial cell expression of Slug[J].J Cell Sci,2014,127(Pt 9):2017-2028.DOI: 10.1242/jcs.143420.
|

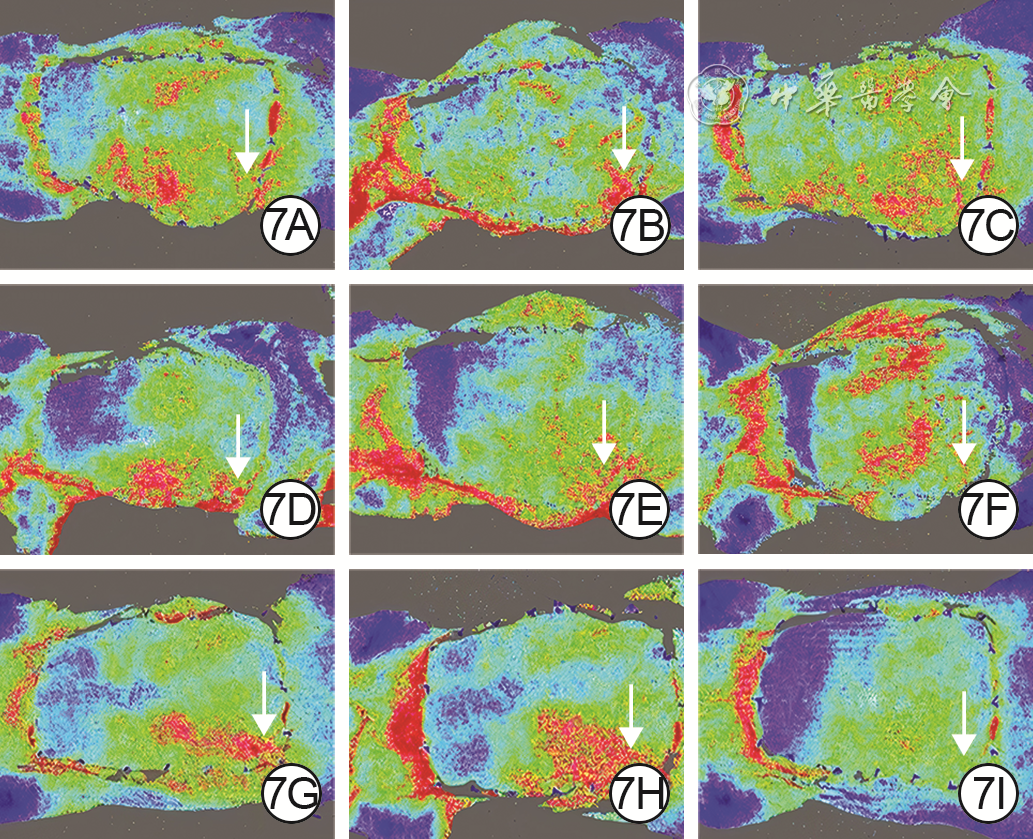
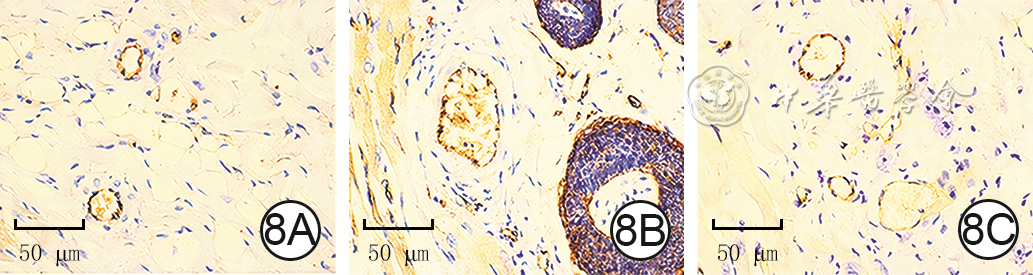
Figures(9) / Tables(3)
Copyright © Chinese Journal of Burns京ICP备07035254号-14
E-mail:shaoshangzazhi@163.com
Supported by:
Beijing Renhe Information Technology Co. Ltd






 DownLoad:
DownLoad: