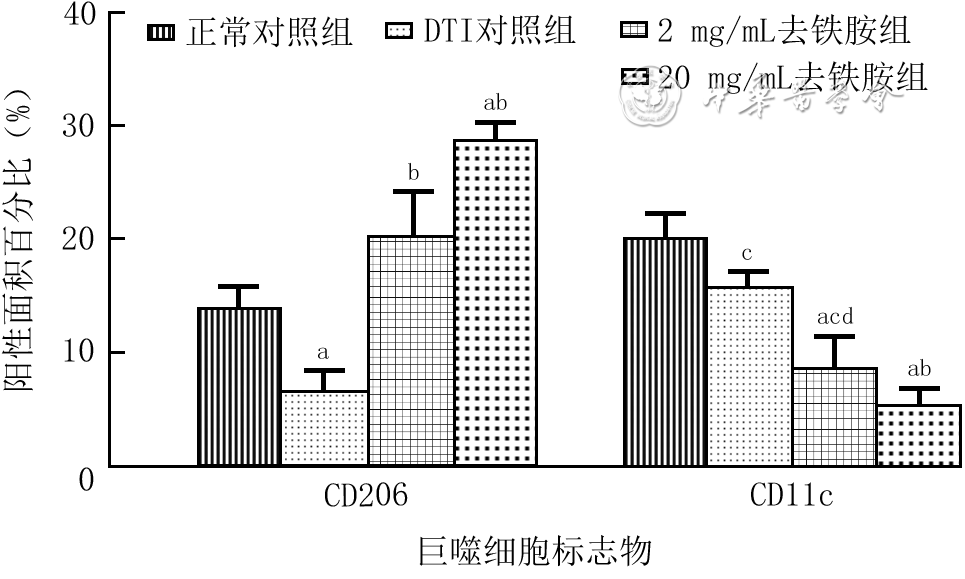
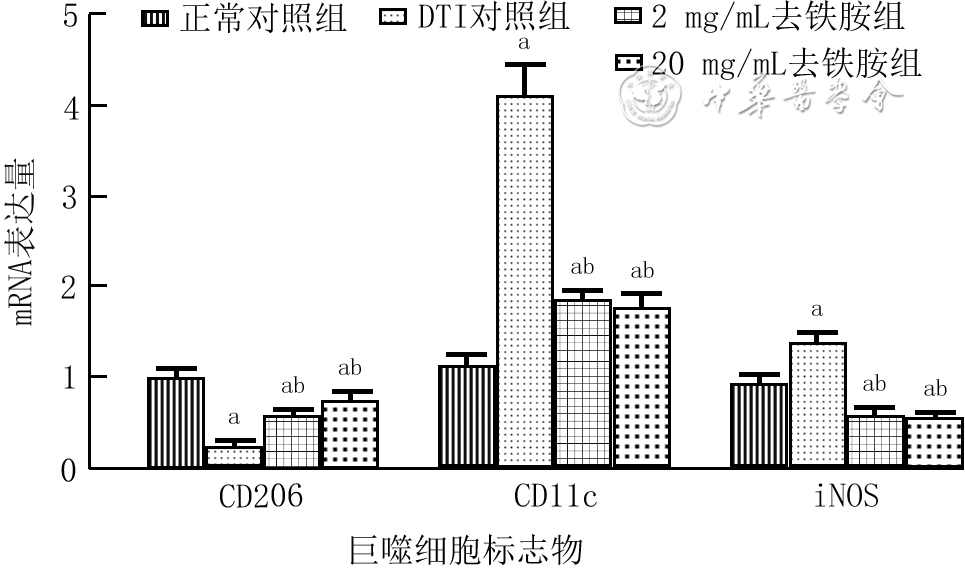
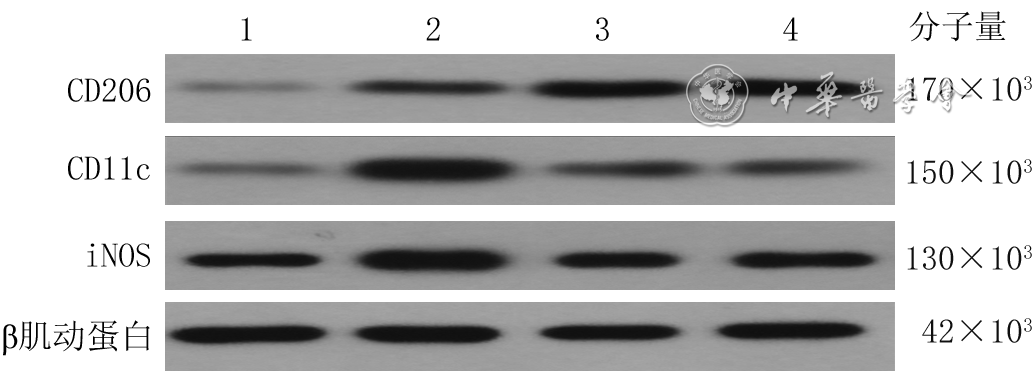
| Citation: | Shan H,Zhang ZR,Wang XY,et al.Regulatory mechanism of deferoxamine on macrophage polarization and wound healing in mice with deep tissue injury[J].Chin J Burns Wounds,2022,38(8):767-777.DOI: 10.3760/cma.j.cn501225-20220114-00007.

|
| [1] |
SenCK. Human wound and its burden: updated 2020 compendium of estimates[J]. Adv Wound Care (New Rochelle), 2021, 10(5):281-292. DOI: 10.1089/wound.2021.0026.
|
| [2] |
FuX. Wound healing center establishment and new technology application in improving the wound healing quality in China[J/OL]. Burns Trauma, 2020, 8: tkaa038[2022-06-06]. https://pubmed.ncbi.nlm.nih.gov/33134399/. DOI: 10.1093/burnst/tkaa038.
|
| [3] |
WynnM. Deep tissue injury: a narrative review on the aetiology of a controversial wound[J]. Br J Nurs, 2021, 30(5):S32-37. DOI: 10.12968/bjon.2021.30.5.S32.
|
| [4] |
ShiH, XieH, ZhaoY, et al. Myoprotective effects of bFGF on skeletal muscle injury in pressure-related deep tissue injury in rats[J/OL]. Burns Trauma, 2016, 4: 26[2022-06-06]. https://pubmed.ncbi.nlm.nih.gov/27574694/.DOI: 10.1186/s41038-016-0051-y.
|
| [5] |
陆树良.把握创面修复的规律和特征促进创面愈合[J].中华烧伤杂志, 2021, 37(5):401-403. DOI: 10.3760/cma.j.cn501120-20210322-00100.
|
| [6] |
WangP, CuiY, RenQ, et al. Mitochondrial ferritin attenuates cerebral ischaemia/reperfusion injury by inhibiting ferroptosis[J]. Cell Death Dis, 2021,12(5):447. DOI: 10.1038/s41419-021-03725-5.
|
| [7] |
KangH, HanM, XueJ, et al. Renal clearable nanochelators for iron overload therapy[J]. Nat Commun, 2019, 10(1):5134. DOI: 10.1038/s41467-019-13143-z.
|
| [8] |
WuS, YangJ, SunG, et al. Macrophage extracellular traps aggravate iron overload-related liver ischaemia/reperfusion injury[J]. Br J Pharmacol, 2021, 178(18):3783-3796. DOI: 10.1111/bph.15518.
|
| [9] |
张子锐, 张亚萍, 郭景琳,等.去铁胺通过抑制氧化应激反应促进小鼠深部组织压力性损伤创面愈合[J].中国病理生理杂志, 2021, 37(9):1646-1654. DOI: 10.3969/j.issn.1000-4718.2021.09.014.
|
| [10] |
SindrilaruA, PetersT, WieschalkaS, et al. An unrestrained proinflammatory M1 macrophage population induced by iron impairs wound healing in humans and mice[J]. J Clin Invest, 2011, 121(3):985-997. DOI: 10.1172/JCI44490.
|
| [11] |
HoldenP, NairLS. Deferoxamine: an angiogenic and antioxidant molecule for tissue regeneration[J]. Tissue Eng Part B Rev, 2019, 25(6):461-470. DOI: 10.1089/ten.TEB.2019.0111.
|
| [12] |
DingJ, WangX, ChenB, et al. Exosomes derived from human bone marrow mesenchymal stem cells stimulated by deferoxamine accelerate cutaneous wound healing by promoting angiogenesis[J]. Biomed Res Int, 2019, 2019:9742765. DOI: 10.1155/2019/9742765.
|
| [13] |
DuscherD, TrotsyukAA, MaanZN, et al. Optimization of transdermal deferoxamine leads to enhanced efficacy in healing skin wounds[J]. J Control Release, 2019, 308:232-239. DOI: 10.1016/j.jconrel.2019.07.009.
|
| [14] |
WinnNC, VolkKM, HastyAH. Regulation of tissue iron homeostasis: the macrophage "ferrostat"[J]. JCI Insight, 2020, 5(2):e132964. DOI: 10.1172/jci.insight.132964.
|
| [15] |
LiuP, YangX, HanJ, et al. Tazarotene-loaded PLGA nanoparticles potentiate deep tissue pressure injury healing via VEGF-Notch signaling[J]. Mater Sci Eng C Mater Biol Appl, 2020, 114:111027. DOI: 10.1016/j.msec.2020.111027.
|
| [16] |
YangX,GuoJL,HanJ,et al.Chitosan hydrogel encapsulated with LL-37 peptide promotes deep tissue injury healing in a mouse model[J].Mil Med Res,2020,7(1):20.DOI: 10.1186/s40779-020-00249-5.
|
| [17] |
SariY,MinematsuT,HuangL,et al.Establishment of a novel rat model for deep tissue injury deterioration[J].Int Wound J,2015,12(2):202-209.DOI: 10.1111/iwj.12082.
|
| [18] |
RavingerováT, KindernayL, BartekováM, et al. The molecular mechanisms of iron metabolism and its role in cardiac dysfunction and cardioprotection[J]. Int J Mol Sci, 2020, 21(21):7889. DOI: 10.3390/ijms21217889.
|
| [19] |
YanHF, TuoQZ, YinQZ, et al. The pathological role of ferroptosis in ischemia/reperfusion-related injury[J]. Zool Res, 2020, 41(3):220-230. DOI: 10.24272/j.issn.2095-8137.2020.042.
|
| [20] |
WilkinsonHN, UpsonSE, BanyardKL, et al. Reduced iron in diabetic wounds: an oxidative stress-dependent role for STEAP3 in extracellular matrix deposition and remodeling[J]. J Invest Dermatol, 2019,139(11):2368-2377.e7. DOI: 10.1016/j.jid.2019.05.014.
|
| [21] |
WangC, CaiY, ZhangY, et al. Local injection of deferoxamine improves neovascularization in ischemic diabetic random flap by increasing HIF-1α and VEGF expression[J]. PLoS One, 2014, 9(6):e100818. DOI: 10.1371/journal.pone.0100818.
|
| [22] |
Tchanque-FossuoCN, DahleSE, BuchmanSR, et al. Deferoxamine: potential novel topical therapeutic for chronic wounds[J]. Br J Dermatol, 2017, 176(4):1056-1059. DOI: 10.1111/bjd.14956.
|
| [23] |
BonhamCA, RodriguesM, GalvezM, et al. Deferoxamine can prevent pressure ulcers and accelerate healing in aged mice[J]. Wound Repair Regen, 2018, 26(3):300-305. DOI: 10.1111/wrr.12667.
|
| [24] |
RamM, SinghV, KumawatS, et al. Deferoxamine modulates cytokines and growth factors to accelerate cutaneous wound healing in diabetic rats[J]. Eur J Pharmacol, 2015, 764:9-21. DOI: 10.1016/j.ejphar.2015.06.029.
|
| [25] |
RenH, ZhaoF, ZhangQ, et al. Autophagy and skin wound healing[J/OL]. Burns Trauma, 2022, 10: tkac003[2022-06-06].https://pubmed.ncbi.nlm.nih.gov/35187180/. DOI: 10.1093/burnst/tkac003.
|
| [26] |
HuP, YangQ, WangQ, et al. Mesenchymal stromal cells-exosomes: a promising cell-free therapeutic tool for wound healing and cutaneous regeneration[J/OL]. Burns Trauma, 2019, 7: 38[2022-06-06]. https://pubmed.ncbi.nlm.nih.gov/31890717/.DOI: 10.1186/s41038-019-0178-8.
|
| [27] |
CaputaG, FlachsmannLJ, CameronAM. Macrophage metabolism: a wound-healing perspective[J]. Immunol Cell Biol, 2019, 97(3):268-278. DOI: 10.1111/imcb.12237.
|
| [28] |
Shapouri-MoghaddamA, MohammadianS, VaziniH, et al. Macrophage plasticity, polarization, and function in health and disease[J]. J Cell Physiol, 2018, 233(9):6425-6440. DOI: 10.1002/jcp.26429.
|
| [29] |
WangS, LiuC, PanS, et al. Deferoxamine attenuates lipopolysaccharide-induced inflammatory responses and protects against endotoxic shock in mice[J]. Biochem Biophys Res Commun, 2015,465(2):305-311. DOI: 10.1016/j.bbrc.2015.08.032.
|
| [30] |
YangF, WuZ, YangD, et al. Characteristics of macrophages from myelodysplastic syndrome microenvironment[J]. Exp Cell Res, 2021, 408(1):112837. DOI: 10.1016/j.yexcr.2021.112837.
|
| [31] |
HamidzadehK, ChristensenSM, DalbyE, et al. Macrophages and the recovery from acute and chronic inflammation[J]. Annu Rev Physiol, 2017, 79:567-592. DOI: 10.1146/annurev-physiol-022516-034348.
|
| [32] |
YangC, HeL, HeP, et al. Increased drug resistance in breast cancer by tumor-associated macrophages through IL-10/STAT3/bcl-2 signaling pathway[J]. Med Oncol, 2015, 32(2):352. DOI: 10.1007/s12032-014-0352-6.
|
| [33] |
TangL, ZhangH, WangC, et al. M2A and M2C macrophage subsets ameliorate inflammation and fibroproliferation in acute lung injury through interleukin 10 pathway[J]. Shock, 2017, 48 (1):119-129. DOI: 10.1097/SHK.0000000000000820.
|
| [34] |
KuangY, GuoW, LingJ, et al. Iron-dependent CDK1 activity promotes lung carcinogenesis via activation of the GP130/STAT3 signaling pathway[J]. Cell Death Dis, 2019, 10(4):297. DOI: 10.1038/s41419-019-1528-y.
|
| [35] |
ZhangZ, ZhangJ, HeP, et al. Interleukin-37 suppresses hepatocellular carcinoma growth through inhibiting M2 polarization of tumor-associated macrophages[J]. Mol Immunol,2020,122:13-20. DOI: 10.1016/j.molimm.2020.03.012.
|
| [36] |
ParkSM, AnJH, LeeJH, et al. Extracellular vesicles derived from DFO-preconditioned canine AT-MSCs reprogram macrophages into M2 phase[J]. PLoS One, 2021, 16(7):e0254657. DOI: 10.1371/journal.pone.0254657.
|
| [37] |
ZhouD, HuangC, LinZ, et al. Macrophage polarization and function with emphasis on the evolving roles of coordinated regulation of cellular signaling pathways[J]. Cell Signal, 2014, 26 (2):192-197. DOI: 10.1016/j.cellsig.2013.11.004.
|
| [38] |
WuWK, LlewellynOP, BatesDO, et al. IL-10 regulation of macrophage VEGF production is dependent on macrophage polarisation and hypoxia[J]. Immunobiology, 2010, 215(9/10):796-803. DOI: 10.1016/j.imbio.2010.05.025.
|
| [39] |
白海良, 段红杰, 陈晨, 等. Janus激酶/信号转导及转录激活子3通路抑制剂对严重烧伤大鼠骨骼肌功能的影响及其机制[J]. 中华烧伤杂志, 2021, 37(3): 271-278. DOI: 10.3760/cma.j.cn501120-20200120-00030.
|
| [40] |
ZhuJ, LuoL, TianL, et al. Aryl hydrocarbon receptor promotes IL-10 expression in inflammatory macrophages through Src-STAT3 signaling pathway[J]. Front Immunol, 2018, 9: 2033. DOI: 10.3389/fimmu.2018.02033.
|


Figures(8) / Tables(4)
Copyright © Chinese Journal of Burns京ICP备07035254号-14
E-mail:shaoshangzazhi@163.com
Supported by:
Beijing Renhe Information Technology Co. Ltd

Pan Bohan, Sun Yu, Tang Tao, et al. Application of antigen-free porcine peritoneum as carrier of autologous microskin grafting in patients with extensive burn[J]. Chin j Burns, 2020, 36(9): 861-864. Doi: 10.3760/cma.j.cn501120-20190725-00311





 DownLoad:
DownLoad: