Liu XX,Liu DW.Research advances on the effects of RNA N6-methyladenosine modification in the relevant pathophysiological processes of wound repair[J].Chin J Burns Wounds,2022,38(10):989-993.DOI: 10.3760/cma.j.cn501120-20210804-00267.
Citation:
Yi JR,Li ZN,Xie HQ,et al.Effects and mechanism of human umbilical vein endothelial cells-derived exosomes on wound healing in diabetic rabbits[J].Chin J Burns Wounds,2022,38(11):1023-1033.DOI: 10.3760/cma.j.cn501225-20220622-00254.
Liu XX,Liu DW.Research advances on the effects of RNA N6-methyladenosine modification in the relevant pathophysiological processes of wound repair[J].Chin J Burns Wounds,2022,38(10):989-993.DOI: 10.3760/cma.j.cn501120-20210804-00267.
Citation:
Yi JR,Li ZN,Xie HQ,et al.Effects and mechanism of human umbilical vein endothelial cells-derived exosomes on wound healing in diabetic rabbits[J].Chin J Burns Wounds,2022,38(11):1023-1033.DOI: 10.3760/cma.j.cn501225-20220622-00254.
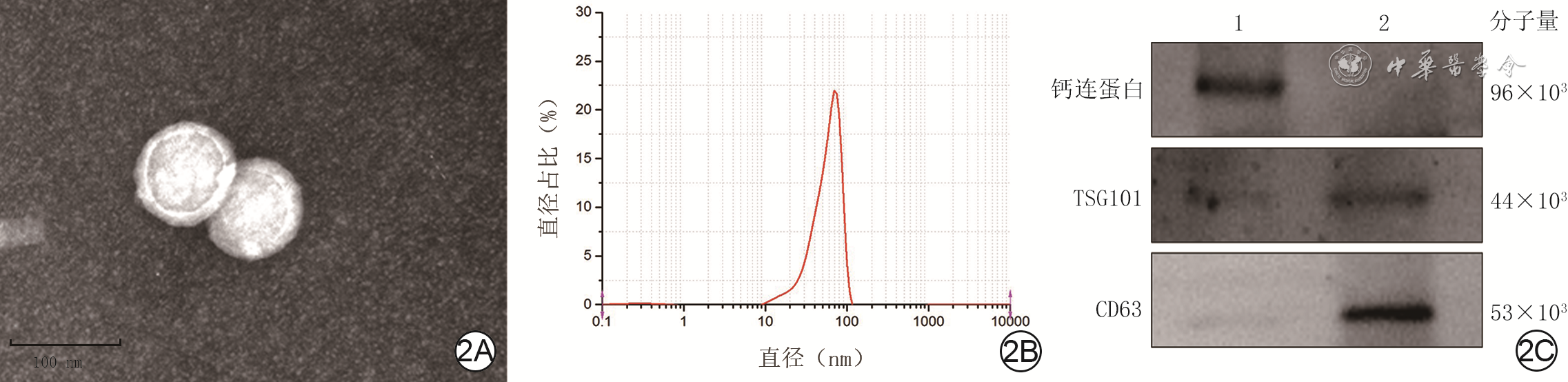
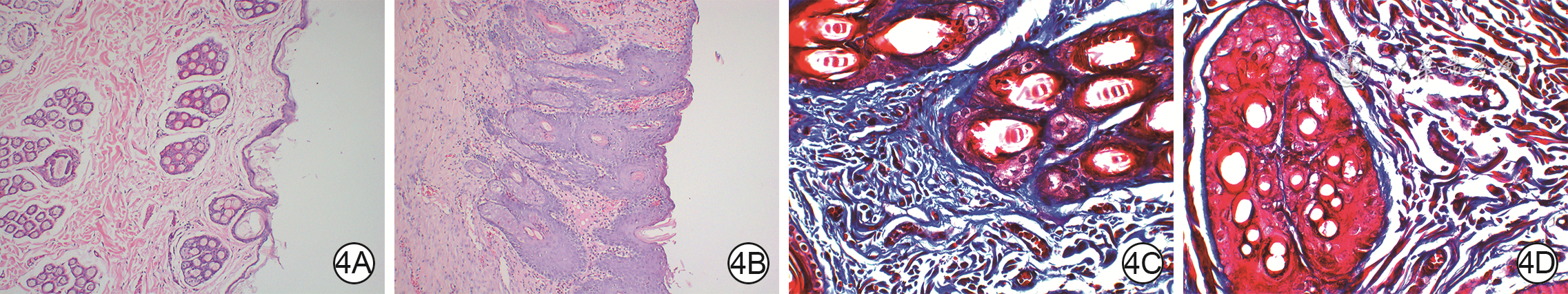
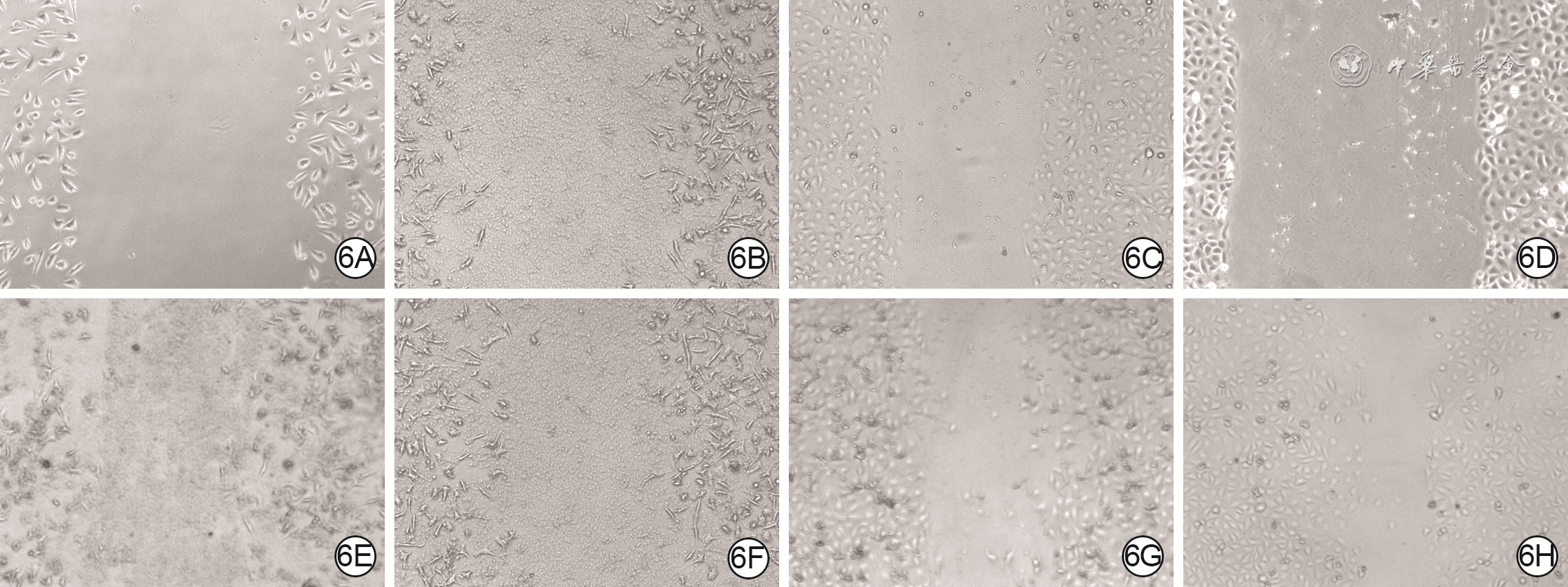
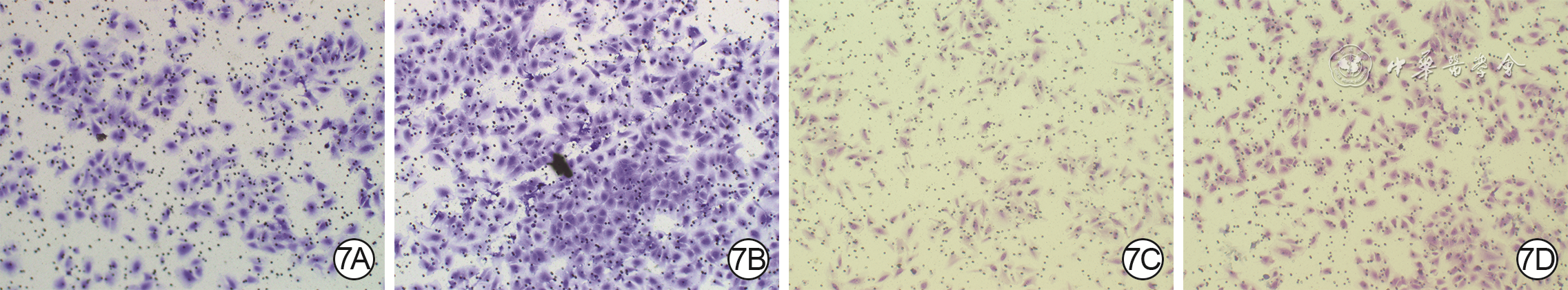
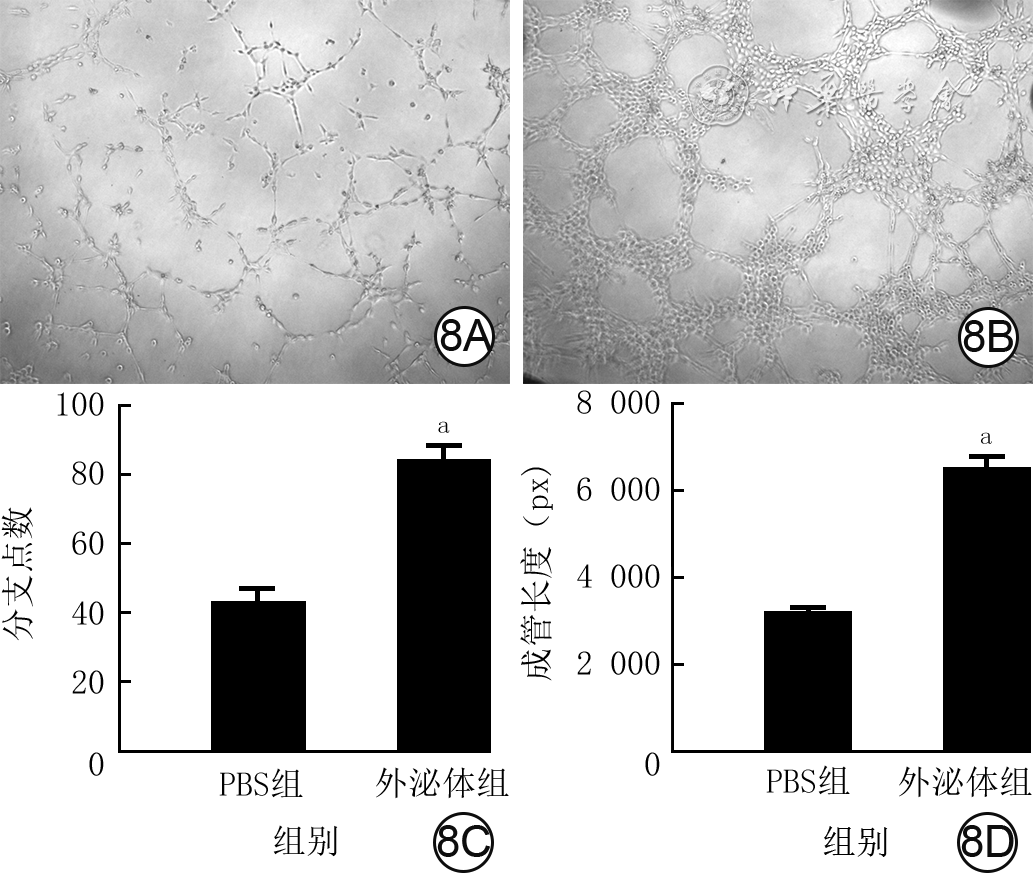
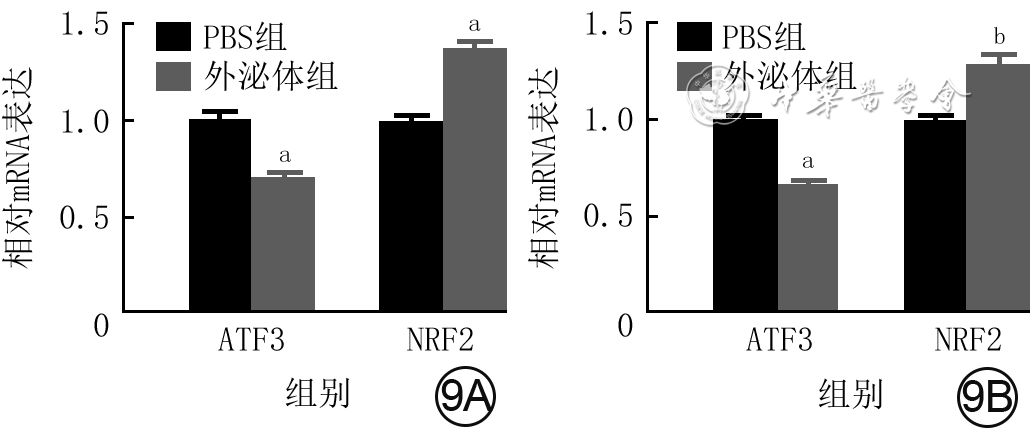
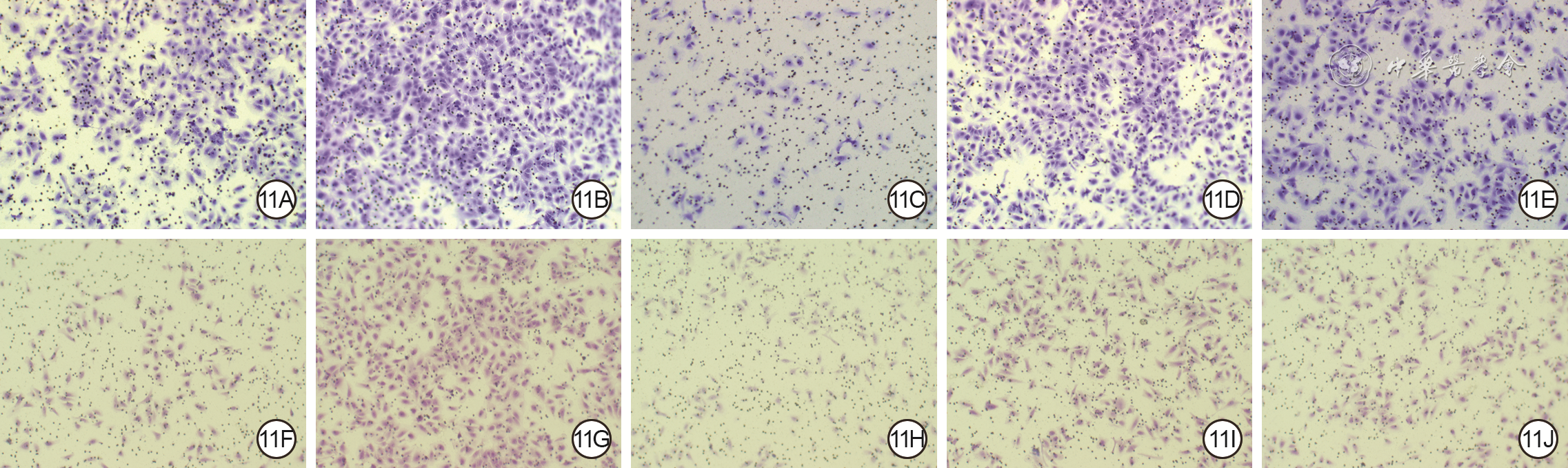
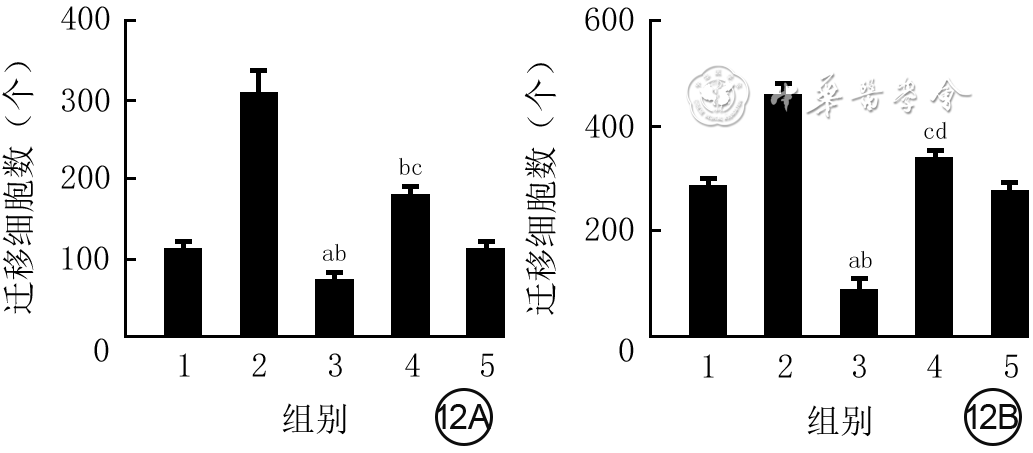
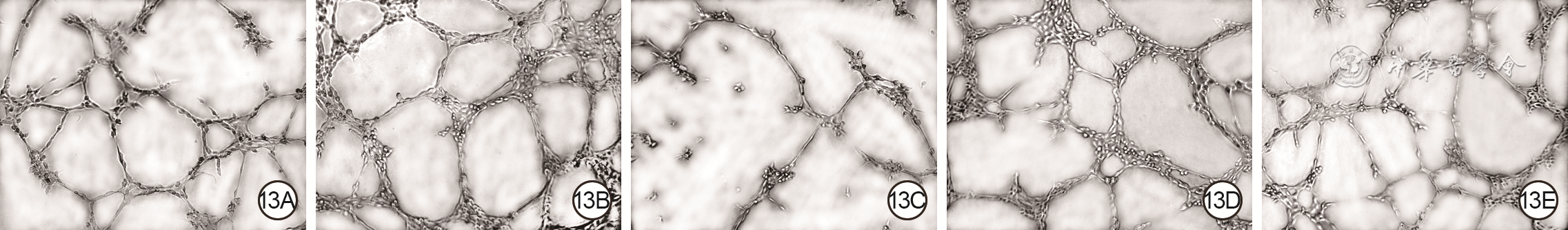
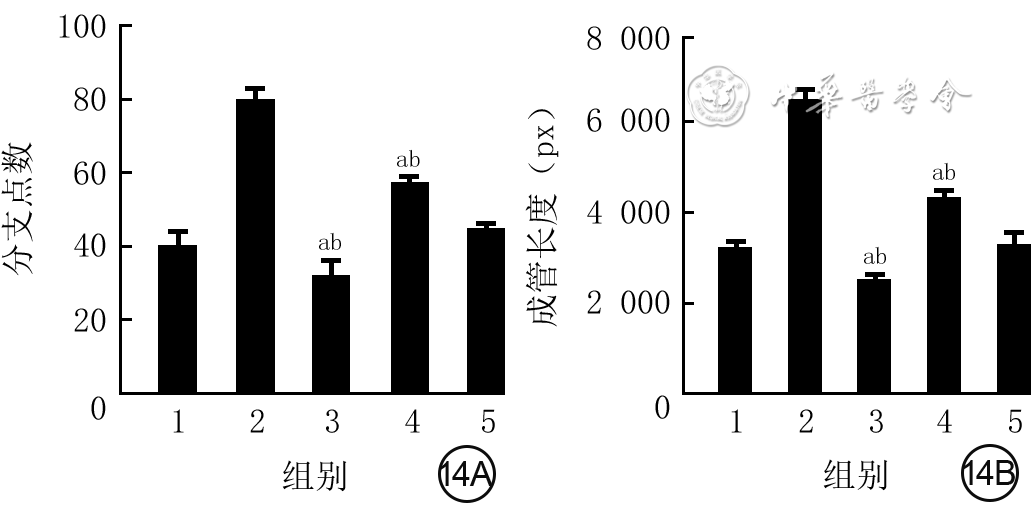
Objective The investigate the effects and mechanism of exosomes derived from human umbilical vein endothelial cells (HUVECs) on wound healing in diabetes rabbits.Methods The experimental research methods were used. The primary vascular endothelial cells (VECs) and human skin fibroblasts (HSFs) were extracted from skin tissue around ulcer by surgical excision of two patients with diabetic ulcer (the male aged 49 years and the female aged 58 years) admitted to Xiangya Third Hospital of Central South University in June 2019. The cells were successfully identified through morphological observation and flow cytometry. The HUVEC exosomes were extracted by ultracentrifugation and identified successfully by morphological observation, particle size detection, and Western blotting detection. Twenty female 3-month-old New Zealand rabbits were taken to create one type 2 diabetic full-thickness skin defect wound respectively on both sides of the back. The wounds were divided into exosomes group and phosphate buffer solution (PBS) group and treated accordingly, with 20 wounds in each group, the time of complete tissue coverage of wound was recorded. On PID 14, hematoxylin-eosin staining or Masson staining was performed to observe angiogenesis or collagen fiber hyperplasia (n=20). The VECs and HSFs were co-cultured with HUVEC exosomes for 24 h to observe the uptake of HUVEC exosomes by the two kinds of cells. The VECs and HSFs were divided to exosome group treated with HUVEC exosomes and PBS group treated with PBS to detect the cell proliferation on 4 d of culture with cell count kit 8, to detect and calculate the cell migration rate at 24 and 48 h after scratch by scratch test, to detect the cell migration number at 24 h of culture with Transwell test, and to detect the mRNA expressions of nuclear factor-erythroid 2-related factor 2 (NRF2) and transcription activating factor 3 (ATF3) by real time fluorescence quantitative reverse transcription polymerase chain reaction. Besides, the number of vascular branches and vascular length were observed in the tube forming experiment after 12 h of culture of VECs (n=3). The VECs and HSFs were taken and divided into PBS group and exosome group treated as before, and NRF2 interference group, ATF3 interference group, and no-load interference group with corresponding gene interference. The proliferation and migration of the two kinds of cells, and angiogenesis of VECs were detected as before (n=3). Data were statistically analyzed with analysis of variance for repeated measurement, one-way analysis of variance, independent sample t test, and least significant difference test.Results The time of complete tissue coverage of wound in exosome group was (17.9±1.9) d, which was significantly shorter than (25.2±2.3) d in PBS group (t=4.54, P<0.05). On PID14, the vascular density of wound in PBS group was significantly lower than that in exosome group (t=10.12, P<0.01), and the collagen fiber hyperplasia was less than that in exosome group. After 24 h of culture, HUVEC exosomes were successfully absorbed by VECs and HSFs. The proliferative activity of HSFs and VECs in exosome group was significantly higher than that in PBS group after 4 d of culture (with t values of 54.73 and 7.05, respectively, P<0.01). At 24 and 48 h after scratch, the migration rates of HSFs (with t values of 3.42 and 11.87, respectively, P<0.05 or P<0.01) and VECs (with t values of 21.42 and 5.49, respectively, P<0.05 or P<0.01) in exosome group were significantly higher than those in PBS group. After 24 h of culture, the migration numbers of VECs and HSFs in exosome group were significantly higher than those in PBS group (with t values of 12.31 and 16.78, respectively, P<0.01). After 12 h of culture, the mRNA expressions of NRF2 in HSFs and VECs in exosome group were significantly higher than those in PBS group (with t values of 7.52 and 5.78, respectively, P<0.05 or P<0.01), and the mRNA expressions of ATF3 were significantly lower than those in PBS group (with t values of 13.44 and 8.99, respectively, P<0.01). After 12 h of culture, the number of vascular branches of VECs in exosome group was significantly more than that in PBS group (t=17.60, P<0.01), and the vascular length was significantly longer than that in PBS group (t=77.30, P<0.01). After 4 d of culture, the proliferation activity of HSFs and VECs in NRF2 interference group was significantly lower than that in PBS group and exosome group (P<0.05 or P<0.01); the proliferation activity of HSFs and VECs in ATF3 interference group was significantly higher than that in PBS group (P<0.05 or P<0.01) and significantly lower than that in exosome group (P<0.05 or P<0.01). At 24 and 48 h after scratch, the migration rates of HSFs and VECs in ATF3 interference group were significantly higher than those in PBS group (P<0.05 or P<0.01) and significantly lower than those in exosome group (P<0.05 or P<0.01). At 24 and 48 h after scratch, the migration rates of HSFs and VECs in NRF2 interference group were significantly lower than those in PBS group and exosome group (P<0.05 or P<0.01). After 24 h of culture, the migration numbers of VECs and HSFs in ATF3 interference group were significantly more than those in PBS group (P<0.05) and significantly less than those in exosome group (P<0.05 or P<0.01); the migration numbers of VECs and HSFs in NRF2 interference group were significantly less than those in PBS group and exosome group (P<0.01). After 12 h of culture, the vascular length and number of branches of VECs in NRF2 interference group were significantly decreased compared with those in PBS group and exosome group (P<0.01); the vascular length and number of branches of VECs in ATF3 interference group were significantly increased compared with those in PBS group (P<0.01) and were significantly decreased compared with those in exosome group (P<0.01).Conclusions HUVEC exosomes can promote the wound healing of diabetic rabbits by promoting the proliferation and migration of VECs and HSFs, and NRF2 and ATF3 are obviously affected by exosomes in this process, which are the possible targets of exosome action.
MezeraV,BurešI.Chronic non-healing wounds in geriatrics[J].Vnitr Lek,2018,64(11):1098-1104.
[2]
HanG,CeilleyR.Chronic wound healing: a review of current management and treatments[J].Adv Ther,2017,34(3):599-610.DOI: 10.1007/s12325-017-0478-y.
[3]
GreeningDW,GopalSK,XuR,et al.Exosomes and their roles in immune regulation and cancer[J].Semin Cell Dev Biol,2015,40:72-81.DOI: 10.1016/j.semcdb.2015.02.009.
[4]
KalaniA,ChaturvediP,KamatPK,et al.Curcumin-loaded embryonic stem cell exosomes restored neurovascular unit following ischemia-reperfusion injury[J].Int J Biochem Cell Biol,2016,79:360-369.DOI: 10.1016/j.biocel.2016.09.002.
[5]
ShabbirA,CoxA,Rodriguez-MenocalL,et al.Mesenchymal stem cell exosomes induce proliferation and migration of normal and chronic wound fibroblasts, and enhance angiogenesis in vitro[J].Stem Cells Dev,2015,24(14):1635-1647.DOI: 10.1089/scd.2014.0316.
CooperDR,WangC,PatelR,et al.Human adipose-derived stem cell conditioned media and exosomes containing MALAT1 promote human dermal fibroblast migration and ischemic wound healing[J].Adv Wound Care (New Rochelle),2018,7(9):299-308.DOI: 10.1089/wound.2017.0775.
[8]
ShafeiS,KhanmohammadiM,HeidariR,et al.Exosome loaded alginate hydrogel promotes tissue regeneration in full-thickness skin wounds: an in vivo study[J].J Biomed Mater Res A,2020,108(3):545-556.DOI: 10.1002/jbm.a.36835.
[9]
WangC,LiangC,WangR,et al.The fabrication of a highly efficient self-healing hydrogel from natural biopolymers loaded with exosomes for the synergistic promotion of severe wound healing[J].Biomater Sci,2019,8(1):313-324.DOI: 10.1039/c9bm01207a.
[10]
YousefiS,LiuT,WangRK.Segmentation and quantification of blood vessels for OCT-based micro-angiograms using hybrid shape/intensity compounding[J].Microvasc Res,2015,97:37-46.DOI: 10.1016/j.mvr.2014.09.007.
[11]
Romana-SouzaB,Assis de BritoTL,PereiraGR,et al.Gonadal hormones differently modulate cutaneous wound healing of chronically stressed mice[J].Brain Behav Immun,2014,36:101-110.DOI: 10.1016/j.bbi.2013.10.015.
[12]
BjørgeIM,KimSY,ManoJF,et al.Extracellular vesicles, exosomes and shedding vesicles in regenerative medicine - a new paradigm for tissue repair[J].Biomater Sci,2017,6(1):60-78.DOI: 10.1039/c7bm00479f.
[13]
KatagiriS,ParkK,MaedaY,et al.Overexpressing IRS1 in endothelial cells enhances angioblast differentiation and wound healing in diabetes and insulin resistance[J].Diabetes,2016,65(9):2760-2771.DOI: 10.2337/db15-1721.
[14]
MurakamiS,MotohashiH.Roles of Nrf2 in cell proliferation and differentiation[J].Free Radic Biol Med,2015,88(Pt B):168-178.DOI: 10.1016/j.freeradbiomed.2015.06.030.
[15]
YiT,LiX,WangE,et al.Activation of the nuclear erythroid 2-related factor 2 antioxidant responsive element (Nrf2-ARE) signaling pathway alleviates acute graft-versus-host disease by reducing oxidative stress and inhibiting infiltration of inflammatory cells in an allogeneic stem cell transplantation mouse model[J].Med Sci Monit,2018,24:5973-5979.DOI: 10.12659/MSM.908130.
[16]
TonelliC,ChioIIC,TuvesonDA.Transcriptional regulation by Nrf2[J].Antioxid Redox Signal,2018,29(17):1727-1745.DOI: 10.1089/ars.2017.7342.
[17]
AmbrozovaN,UlrichovaJ,GalandakovaA.Models for the study of skin wound healing. The role of Nrf2 and NF-κB[J].Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub,2017,161(1):1-13.DOI: 10.5507/bp.2016.063.
[18]
LabzinLI,SchmidtSV,MastersSL,et al.ATF3 is a key regulator of macrophage IFN responses[J].J Immunol,2015,195(9):4446-4455.DOI: 10.4049/jimmunol.1500204.
[19]
ShaH,ZhangD,ZhangY,et al.ATF3 promotes migration and M1/M2 polarization of macrophages by activating tenascin-C via Wnt/β-catenin pathway[J].Mol Med Rep,2017,16(3):3641-3647.DOI: 10.3892/mmr.2017.6992.
[20]
HuY,RaoSS,WangZX,et al.Exosomes from human umbilical cord blood accelerate cutaneous wound healing through miR-21-3p-mediated promotion of angiogenesis and fibroblast function[J].Theranostics,2018,8(1):169-184.DOI: 10.7150/thno.21234.
[21]
MichalikKM,YouX,ManavskiY,et al.Long noncoding RNA MALAT1 regulates endothelial cell function and vessel growth[J].Circ Res,2014,114(9):1389-1397.DOI: 10.1161/CIRCRESAHA.114.303265.
[22]
AlamandaV,SinghS,LawrenceNJ,et al.Nicotine-mediated induction of E-selectin in aortic endothelial cells requires Src kinase and E2F1 transcriptional activity[J].Biochem Biophys Res Commun,2012,418(1):56-61.DOI: 10.1016/j.bbrc.2011.12.127.
[23]
WuX,ShowiheenS,SunAR,et al.Exosomes extraction and identification[J].Methods Mol Biol,2019,2054:81-91.DOI: 10.1007/978-1-4939-9769-5_4.
[24]
TongJ,YangF,LiX,et al.Mechanical characterization and material modeling of diabetic aortas in a rabbit model[J].Ann Biomed Eng,2018,46(3):429-442.DOI: 10.1007/s10439-017-1955-9.
[25]
SedovaKA,AzarovJE,ArteyevaNV,et al.Mechanism of electrocardiographic T-wave flattening in diabetes mellitus: experimental and simulation study[J].Physiol Res,2017,66(5):781-789.DOI: 10.33549/physiolres.933494.
[26]
ZhangY,BiJ,HuangJ,et al.Exosome: a review of its classification, isolation techniques, storage, diagnostic and targeted therapy applications[J].Int J Nanomedicine,2020,15:6917-6934.DOI: 10.2147/IJN.S264498.
[27]
WeiP, ZhongC, YangX, et al. Exosomes derived from human amniotic epithelial cells accelerate diabetic wound healing via PI3K-AKT-mTOR-mediated promotion in angiogenesis and fibroblast function[J/OL]. Burns Trauma, 2020,8:tkaa020[2022-10-31]. https://pubmed.ncbi.nlm.nih.gov/32923490/.DOI: 10.1093/burnst/tkaa020.
TangPM,ZhouS,MengXM,et al.Smad3 promotes cancer progression by inhibiting E4BP4-mediated NK cell development[J].Nat Commun,2017,8:14677.DOI: 10.1038/ncomms14677.
[30]
GonçalvesLR,de Oliveira da Silva B,ScarinciLD,et al.MicroRNA-1254 contributes to the controlling of pro-fibrogenic environment in LX-2 cells by modulating SMAD3 and wound repair: new insights in hepatic fibrosis[J].Cell Biol Int,2019,43(3):333-343.DOI: 10.1002/cbin.11075.
[31]
ChenC, GeC, LiuZ,et al. ATF3 inhibits the tumorigenesis and progression of hepatocellular carcinoma cells via upregulation of CYR61 expression[J]. J Exp Clin Cancer Res,2018,37(1):263. DOI: 10.1186/s13046-018-0919-8.
Liu XX,Liu DW.Research advances on the effects of RNA N6-methyladenosine modification in the relevant pathophysiological processes of wound repair[J].Chin J Burns Wounds,2022,38(10):989-993.DOI: 10.3760/cma.j.cn501120-20210804-00267.
Liu XX,Liu DW.Research advances on the effects of RNA N6-methyladenosine modification in the relevant pathophysiological processes of wound repair[J].Chin J Burns Wounds,2022,38(10):989-993.DOI: 10.3760/cma.j.cn501120-20210804-00267.





 下载:
下载:





 DownLoad:
DownLoad: