Lou JQ,Li Y,Cui QW,et al.A prospectively randomized controlled study of the effects of intensive insulin therapy combined with glutamine on nutritional metabolism, inflammatory response, and hemodynamics in severe burn patients[J].Chin J Burns,2021,37(9):821-830.DOI: 10.3760/cma.j.cn501120-20210428-00159.
Citation:
Zhang YP,Zhang Q,Deng F,et al.Effect of P62 on the migration and motility of human epidermal cell line HaCaT in high glucose microenvironment and its mechanism[J].Chin J Burns Wounds,2022,38(11):1014-1022.DOI: 10.3760/cma.j.cn501225-20220630-00272.
Lou JQ,Li Y,Cui QW,et al.A prospectively randomized controlled study of the effects of intensive insulin therapy combined with glutamine on nutritional metabolism, inflammatory response, and hemodynamics in severe burn patients[J].Chin J Burns,2021,37(9):821-830.DOI: 10.3760/cma.j.cn501120-20210428-00159.
Citation:
Zhang YP,Zhang Q,Deng F,et al.Effect of P62 on the migration and motility of human epidermal cell line HaCaT in high glucose microenvironment and its mechanism[J].Chin J Burns Wounds,2022,38(11):1014-1022.DOI: 10.3760/cma.j.cn501225-20220630-00272.
Department of Endocrinology, the First Affiliated Hospital of Army Medical University (the Third Military Medical University), Chongqing 400038, China
2.
State Key Laboratory of Trauma, Burns and Combined Injury, Institute of Burn Research, the First Affiliated Hospital of Army Medical University (the Third Military Medical University), Chongqing 400038, China
3.
Department of Geriatric Oncology, Department of Palliative Care, Chongqing University Cancer Hospital, Chongqing 400030, China
Funds:
Youth Science Foundation Project of National Natural Science Foundation of China82100889
Chongqing Doctor "Through Train" ProjectCSTB2022BSXM-JCX0022
Science and Technology Ability Promotion Project of Army Medical UniversityXZ-2019-505-018
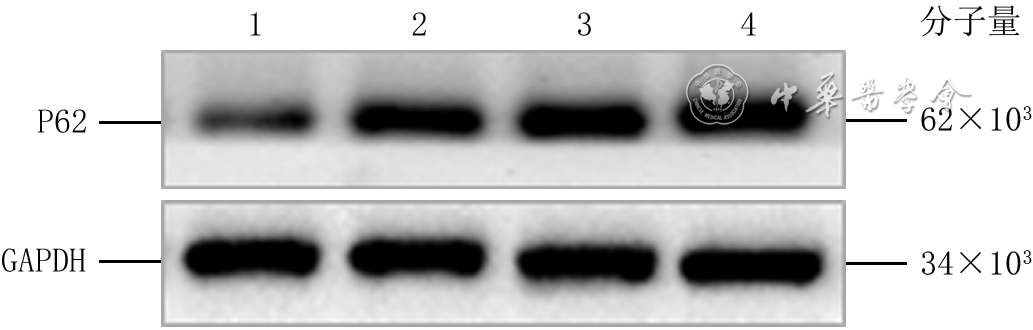
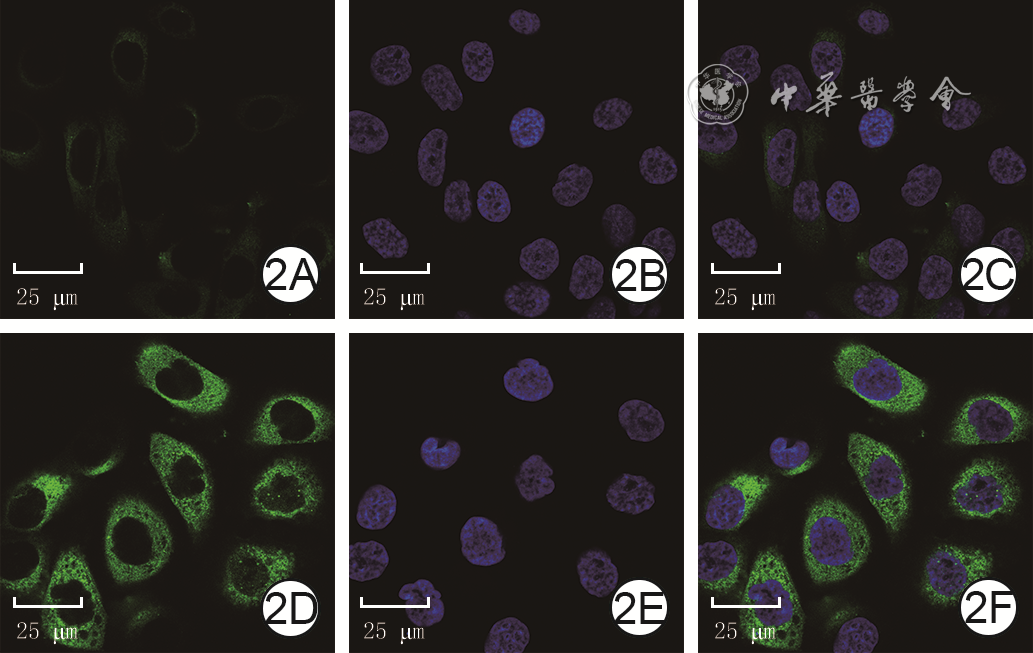
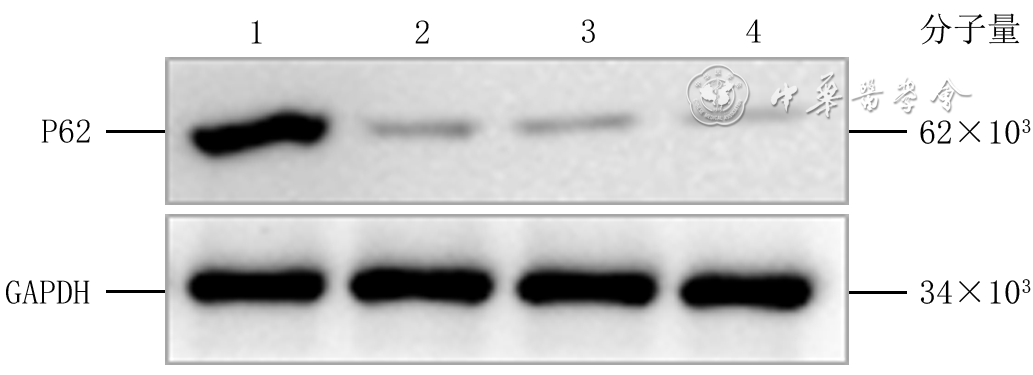
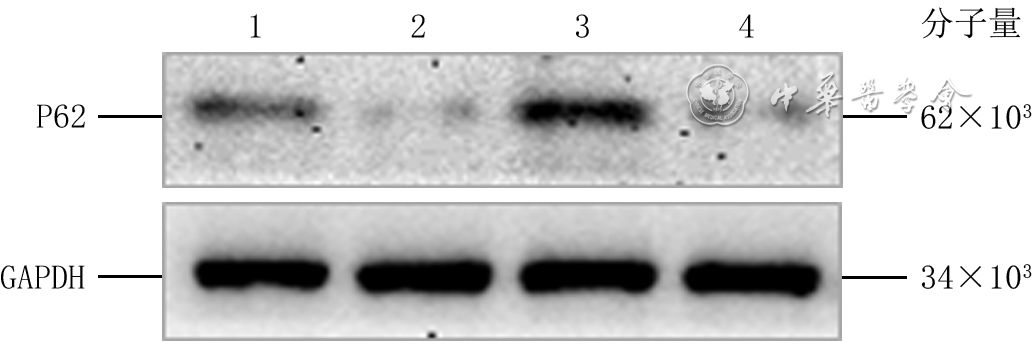
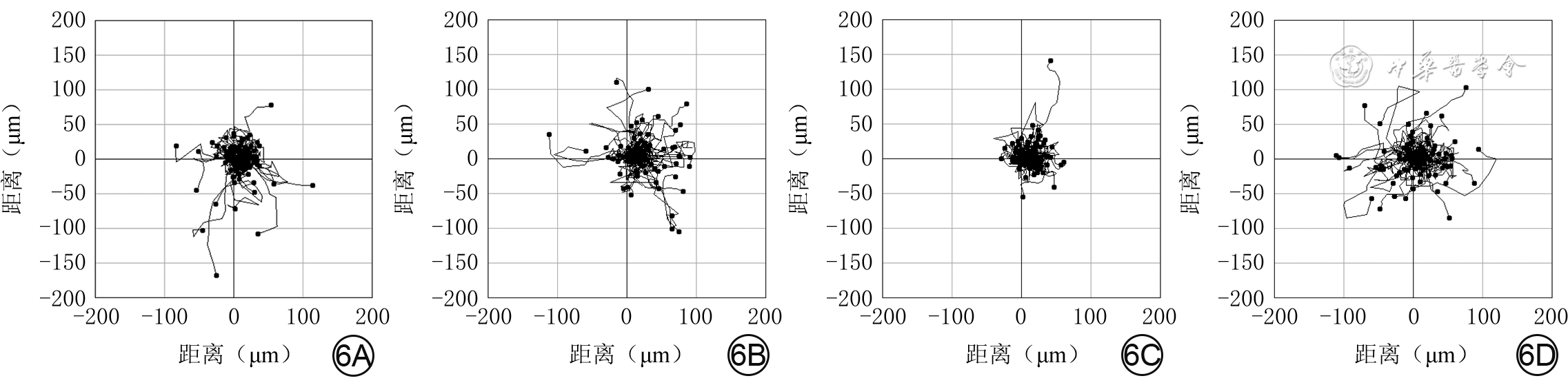
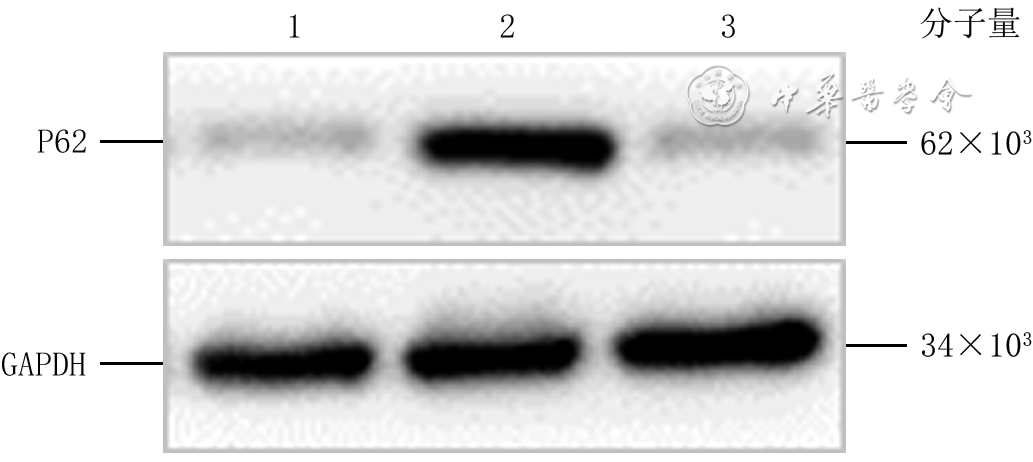
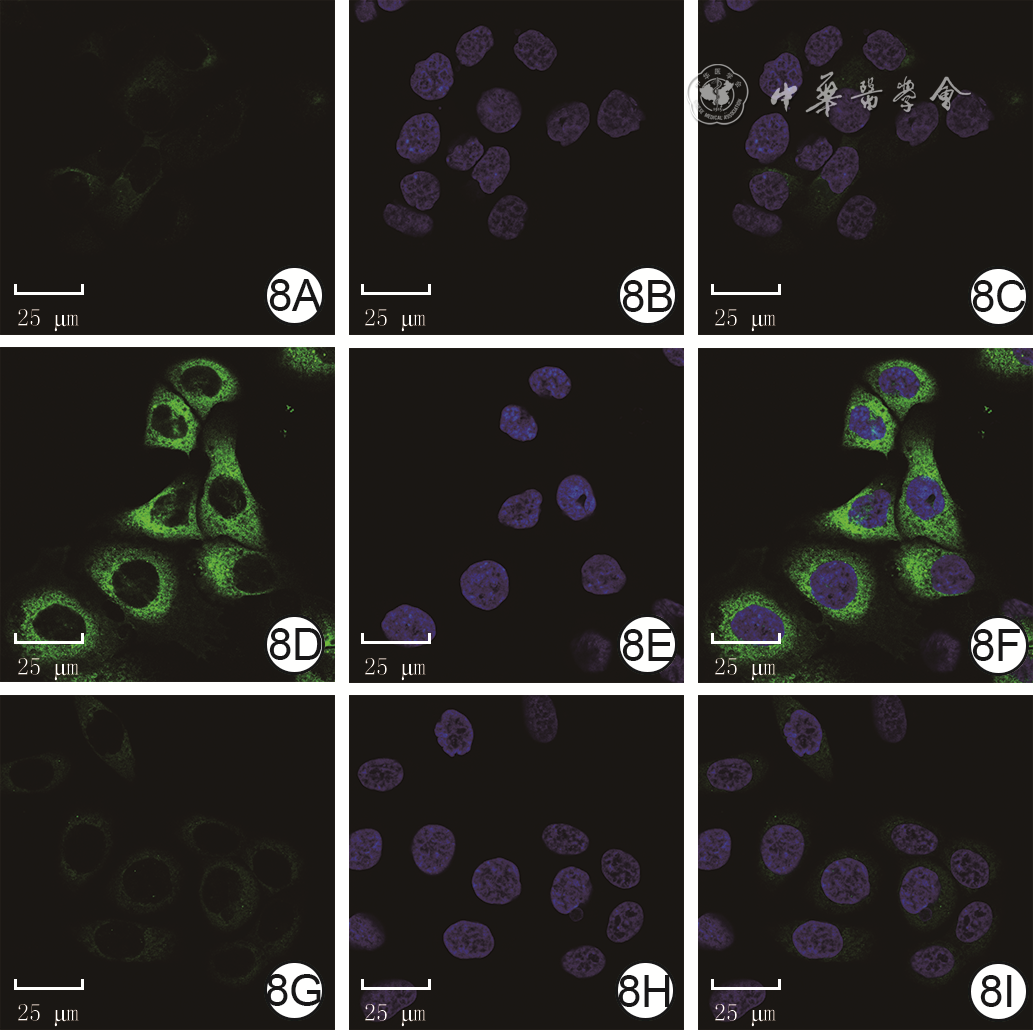
Objective To investigate the effect of P62 on the migration and motility of human epidermal cell line HaCaT in high glucose microenvironment and its possible molecular mechanism, so as to explore the mechanism of refractory diabetic foot wound healing.Methods The method of experimental research was used. HaCaT cells in logarithmic growth phase was taken for experiment. The cells were collected and divided into normal control group (culture solution containing glucose with final molarity of 5.5 mmol/L) and high glucose (culture solution containing glucose with final molarity of 30.0 mmol/L) 24 h group, high glucose 48 h group, and high glucose 72 h group according to the random number table (the same grouping method below). The cells in normal control group were routinely cultured for 72 h, cells in high glucose 72 h group were cultured with high glucose for 72 h, cells in high glucose 48 h group were routinely cultured for 24 h then cultured with high glucose for 48 h, cells in high glucose 24 h group were routinely cultured for 48 h then cultured with high glucose for 24 h. Then the protein expression of P62 was detected by Western blotting. The cells were collected and divided into normal control group and high glucose group. After being correspondingly cultured for 48 h as before, the protein expression of P62 was detected by immunofluorescence method (indicated as green fluorescence). The cells were collected and divided into negative control small interfering RNA (siRNA) group, P62-siRNA-1 group, P62-siRNA-2 group, and P62-siRNA-3 group, and transfected with the corresponding reagents. At post transfection hour (PTH) 72, the protein expression of P62 was detected by Western blotting. The cells were collected and divided into normal glucose+negative control siRNA group, normal glucose+P62-siRNA group, high glucose+negative control siRNA group, and high glucose+P62-siRNA group. After the corresponding treatment, the protein expression of P62 was detected by Western blotting at PTH 72 h, the cell migration rate was detected and calculated at 24 h after scratching by scratch test, with the number of samples being 9; and the range of cell movement was observed and the trajectory velocity was calculated within 3 h under the living cell workstation, with the number of samples being 76, 75, 80, and 79 in normal glucose+negative control siRNA group, normal glucose+P62-siRNA group, high glucose+negative control siRNA group, and high glucose+P62-siRNA group, respectively. The cells were collected and divided into normal glucose+phosphate buffered solution (PBS) group, high glucose+PBS group, and high glucose+N-acetylcysteine (NAC) group. After the corresponding treatment, the protein expression of P62 at 48 h of culture was detected by Western blotting and immunofluorescence method, respectively. Except for scratch test and cell motility experiment, the number of samples was all 3 in the rest experiments. Data were statistically analyzed with one-way analysis of variance and least significant difference test.Results Compared with the protein expression in normal control group, the protein expressions of P62 of cells in high glucose 24 h group, high glucose 48 h group, and high glucose 72 h group were significantly increased (P<0.01). At 48 h of culture, the green fluorescence of P62 of cells in high glucose group was stronger than that in normal control group. At PTH 72, compared with the protein expression in negative control siRNA group, the protein expressions of P62 of cells in P62-siRNA-1 group, P62-siRNA-2 group, and P62-siRNA-3 group were significantly decreased (P<0.01). At PTH 72, compared with the protein expression in normal glucose+negative control siRNA group, the protein expression of P62 of cells in normal glucose+P62-siRNA group was significantly decreased (P<0.01), while the protein expression of P62 of cells in high glucose+negative control siRNA group was significantly increased (P<0.01); compared with the protein expression in high glucose+negative control siRNA group, the protein expression of P62 of cells in high glucose+P62-siRNA group was significantly decreased (P<0.01). At 24 h after scratching, compared with (55±7)% in normal glucose+negative control siRNA group, the cell migration rate in normal glucose+P62-siRNA group was significantly increased ((72±14)%, P<0.01), while the cell migration rate in high glucose+negative control siRNA group was significantly decreased ((37±7)%, P<0.01); compared with that in high glucose+negative control siRNA group, the cell migration rate in high glucose+P62-siRNA group was significantly increased ((54±10)%, P<0.01). Within 3 h of observation, the cell movement range in high glucose+negative control siRNA group was smaller than that in normal glucose+negative control siRNA group, while the cell movement range in normal glucose+P62-siRNA group was larger than that in normal glucose+negative control siRNA group, and the cell movement range in high glucose+P62-siRNA group was larger than that in high glucose+negative control siRNA group. Compared with that in normal glucose+negative control siRNA group, the cell trajectory speed in normal glucose+P62-siRNA group was significantly increased (P<0.01), while the cell trajectory speed in high glucose+negative control siRNA group was significantly decreased (P<0.01); compared with that in high glucose+negative control siRNA group, the cell trajectory speed in high glucose+P62-siRNA group was significantly increased (P<0.01). At 48 h of culture, compared with that in normal glucose+PBS group, the protein expression of P62 of cells in high glucose+PBS group was significantly increased (P<0.01); compared with that in high glucose+PBS group, the protein expression of P62 of cells in high glucose+NAC group was significantly decreased (P<0.01). At 48 h of culture, the green fluorescence of P62 of cells in high glucose+PBS group was stronger than that in normal glucose+PBS group, while the green fluorescence of P62 of cells in high glucose+NAC group was weaker than that in high glucose+PBS group.Conclusions In HaCaT cells, high glucose microenvironment can promote the protein expression of P62; knockdown of P62 protein can promote the migration and increase the mobility of HaCaT cells; and the increase of reactive oxygen species in high glucose microenvironment may be the underlying mechanism for the increase of P62 expression.
(1)分别于治疗1、3、7、14 d 8:00抽取4组患者静脉血5 mL,送至徐州医科大学附属淮海医院检验科检测血糖、白蛋白、前白蛋白、白细胞计数、降钙素原和C反应蛋白(CRP)。(2)每日取3次PiCCO监测参数的平均值为最终结果。统计4组患者治疗1、3、7 d心脏指数、每搏量指数(SVI)、全心舒张末期容积指数(GEDVI)、外周血管阻力指数(SVRI)、血管外肺水指数(EVLWI)、肺血管通透性指数(PVPI)。
ShiMQ, DuZ, QiYC, et al. Wound microenvironment-responsive glucose consumption and hydrogen peroxide generation synergistic with azithromycin for diabetic wounds healing[J]. Theranostics, 2022, 12(6): 2658-2673. DOI: 10.7150/thno.64244.
[4]
KlionskyDJ, PetroniG, AmaravadiRK, et al. Autophagy in major human diseases[J]. EMBO J, 2021, 40(19): e108863. DOI: 10.15252/embj.2021108863.
[5]
RenHY, ZhaoF, ZhangQQ, et al. Autophagy and skin wound healing[J/OL]. Burns Trauma, 2022, 10:tkac003[2022-06-30]. https://pubmed.ncbi.nlm.nih.gov/35187180/.DOI: 10.1093/burnst/tkac003.
[6]
LevineB, KroemerG. Biological functions of autophagy genes: a disease perspective[J]. Cell, 2019, 176(1/2): 11-42. DOI: 10.1016/j.cell.2018.09.048.
[7]
FanXL, HuangTT, TongYD, et al. p62 works as a hub modulation in the ageing process[J]. Ageing Res Rev, 2022, 73: 101538.DOI: 10.1016/j.arr.2021.101538.
[8]
ChaoXJ,NiHM,DingWX. An unexpected tumor suppressor role of SQSTM1/p62 in liver tumorigenesis[J].Autophagy,2022,18(2):459-461.DOI: 10.1080/15548627.2021.2008693.
[9]
KarrasP,Riveiro-FalkenbachE,CañónE,et al.p62/SQSTM1 fuels melanoma progression by opposing mRNA decay of a selective set of pro-metastatic factors[J].Cancer Cell,2019,35(1):46-63.e10.DOI: 10.1016/j.ccell.2018.11.008.
[10]
Serrano-SaenzS, PalaciosC, Delgado-BellidoD, et al. PIM kinases mediate resistance of glioblastoma cells to TRAIL by a p62/SQSTM1-dependent mechanism[J]. Cell Death Dis, 2019, 10(2): 51. DOI: 10.1038/s41419-018-1293-3.
[11]
IslamMA,SooroMA,ZhangPH.Autophagic regulation of p62 is critical for cancer therapy[J].Int J Mol Sci,2018,19(5):1405.DOI: 10.3390/ijms19051405.
[12]
Sánchez-MartínP,KomatsuM.p62/SQSTM1-steering the cell through health and disease[J].J Cell Sci,2018,131(21): jcs.222836.DOI: 10.1242/jcs.222836.
[13]
MildenbergerJ, JohanssonI, SerginI, et al. N-3 PUFAs induce inflammatory tolerance by formation of KEAP1-containing SQSTM1/p62-bodies and activation of NFE2L2[J]. Autophagy, 2017, 13(10): 1664-1678.DOI: 10.1080/15548627.2017.1345411.
[14]
LiLF, ZhangJH, ZhangQ, et al. High glucose suppresses keratinocyte migration through the inhibition of p38 MAPK/autophagy pathway[J]. Front Physiol, 2019, 10: 24. DOI: 10.3389/fphys.2019.00024.
[15]
ZhangJH,LiLF,ZhangQ,et al.Microtubule-associated protein 4 phosphorylation regulates epidermal keratinocyte migration and proliferation[J].Int J Biol Sci,2019,15(9):1962-1976.DOI: 10.7150/ijbs.35440.
[16]
JiangT,LiQY,QiuJM,et al.Nanobiotechnology: applications in chronic wound healing[J].Int J Nanomedicine,2022,17:3125-3145.DOI: 10.2147/IJN.S372211.
[17]
ZhangJH, ZhangC, JiangXP, et al. Involvement of autophagy in hypoxia-BNIP3 signaling to promote epidermal keratinocyte migration[J]. Cell Death Dis, 2019, 10(3): 234. DOI: 10.1038/s41419-019-1473-9.
[18]
LiBM, TangHW, BianXW, et al. Calcium silicate accelerates cutaneous wound healing with enhanced re-epithelialization through EGF/EGFR/ERK-mediated promotion of epidermal stem cell functions[J/OL]. Burns Trauma, 2021, 9: tkab029[2022-06-30]. https://pubmed.ncbi.nlm.nih.gov/34604395/.DOI: 10.1093/burnst/tkab029.
[19]
ShawTJ,MartinP.Wound repair: a showcase for cell plasticity and migration[J].Curr Opin Cell Biol,2016,42:29-37.DOI: 10.1016/j.ceb.2016.04.001.
[20]
HuSC,LanCE.High-glucose environment disturbs the physiologic functions of keratinocytes: focusing on diabetic wound healing[J].J Dermatol Sci,2016,84(2):121-127.DOI: 10.1016/j.jdermsci.2016.07.008.
[21]
ParmarUM, JalgaonkarMP, KulkarniYA, et al. Autophagy-nutrient sensing pathways in diabetic complications[J]. Pharmacol Res, 2022, 184: 106408. DOI: 10.1016/j.phrs.202-2.106408.
NingSB, WangL. The multifunctional protein p62 and its mechanistic roles in cancers[J]. Curr Cancer Drug Targets, 2019, 19(6): 468-478. DOI: 10.2174/1568009618666181016164920.
[24]
TaoMM, LiuT, YouQD, et al. p62 as a therapeutic target for tumor[J]. Eur J Med Chem, 2020, 193: 112231. DOI: 10.1016/j.ejmech.2020.112231.
[25]
ZhangZ,CostaM.p62 functions as a signal hub in metal carcinogenesis[J].Semin Cancer Biol,2021,76:267-278.DOI: 10.1016/j.semcancer.2021.04.014.
[26]
YouZY,JiangWX,QinLY,et al.Requirement for p62 acetylation in the aggregation of ubiquitylated proteins under nutrient stress[J].Nat Commun,2019,10(1):5792.DOI: 10.1038/s41467-019-13718-w.
[27]
GradaA, Otero-VinasM, Prieto-CastrilloF, et al. Research techniques made simple: analysis of collective cell migration using the wound healing assay[J]. J Invest Dermatol, 2017, 137(2): e11-e16.DOI: 10.1016/j.jid.2016.11.020.
DengLL, DuCZ, SongPY, et al. The role of oxidative stress and antioxidants in diabetic wound healing[J]. Oxid Med Cell Longev, 2021, 2021: 8852759. DOI: 10.1155/2021/8852759.
[30]
WeiXR, LiMX, ZhengZJ, et al. Senescence in chronic wounds and potential targeted therapies[J/OL]. Burns Trauma, 2022, 10: tkab045[2022-06-30]. https://pubmed.ncbi.nlm.nih.gov/35187179/.DOI: 10.1093/burnst/tkab045.
[31]
QiuJX, ZhangT, ZhuXY, et al. Hyperoside induces breast cancer cells apoptosis via ROS-mediated NF-κB signaling pathway[J]. Int J Mol Sci, 2020, 21(1): 131. DOI: 10.3390/ijms21010131.
ShenY,YangJJ,ZhaoJ,et al.The switch from ER stress-induced apoptosis to autophagy via ROS-mediated JNK/p62 signals: a survival mechanism in methotrexate-resistant choriocarcinoma cells[J].Exp Cell Res,2015,334(2):207-218.DOI: 10.1016/j.yexcr.2015.04.010.
[34]
BalszuweitF, MenacherG, SchmidtA, et al. Protective effects of the thiol compounds GSH and NAC against sulfur mustard toxicity in a human keratinocyte cell line[J]. Toxicol Lett, 2016, 244: 35-43. DOI: 10.1016/j.toxlet.2015.09.002.
[35]
RizwanH, PalS, SabnamS, et al. High glucose augments ROS generation regulates mitochondrial dysfunction and apoptosis via stress signalling cascades in keratinocytes[J]. Life Sci, 2020, 241: 117148. DOI: 10.1016/j.lfs.2019.117148.
Lou JQ,Li Y,Cui QW,et al.A prospectively randomized controlled study of the effects of intensive insulin therapy combined with glutamine on nutritional metabolism, inflammatory response, and hemodynamics in severe burn patients[J].Chin J Burns,2021,37(9):821-830.DOI: 10.3760/cma.j.cn501120-20210428-00159.
Lou JQ,Li Y,Cui QW,et al.A prospectively randomized controlled study of the effects of intensive insulin therapy combined with glutamine on nutritional metabolism, inflammatory response, and hemodynamics in severe burn patients[J].Chin J Burns,2021,37(9):821-830.DOI: 10.3760/cma.j.cn501120-20210428-00159.




 下载:
下载:





 DownLoad:
DownLoad: