




Two-step two-sample mediation MR analysis of causal relationships between human gut microbiota features, immune cell phenotypes, and HS
-
摘要:
目的 探讨人肠道菌群特征和免疫细胞表型与增生性瘢痕(HS)之间的因果关系。 方法 该研究为基于两步双样本中介孟德尔随机化(MR)分析的研究。从全基因组关联分析数据库中获取人肠道菌群特征、免疫细胞表型、HS的数据,采用逆方差加权法评估119个肠道菌群特征、731种免疫细胞表型与HS之间的因果关系,并通过Cochran Q检验、MR-Egger回归检验分别评估前述关联的异质性与水平多效性,采用两步MR量化免疫细胞表型在肠道菌群特征与HS关联中的中介效应。 结果 7种肠道菌群特征与HS形成风险存在显著因果关系,其中,放线菌门-放线菌纲-双歧杆菌目、放线菌门-放线菌纲-双歧杆菌目-双歧杆菌科、拟杆菌门-拟杆菌纲-拟杆菌目-理研菌科-阿尔斯提普斯属-塞内加尔阿尔斯提普斯种、厚壁菌门-梭菌纲-梭菌目-梭菌科、甘氨酸起始的血红素生物合成超途径、含内消旋二氨基庚二酸的肽聚糖生物合成Ⅰ与HS形成风险均呈显著负相关(OR分别为0.804、0.804、0.784、0.820、0.864、0.686,95%CI分别为0.649~0.996、0.649~0.996、0.623~0.988、0.687~0.980、0.759~0.984、0.491~0.959,P<0.05),厚壁菌门-梭菌纲-梭菌目-真杆菌科-真杆菌属-纤毛真杆菌种与HS形成风险呈显著正相关(OR=1.239,95%CI为1.007~1.525,P<0.05);23种免疫细胞表型与HS形成风险存在显著因果关系,其中,IgD⁻CD38⁻B细胞占B细胞的百分比、CD11c⁺人类白细胞抗原-DR(HLA-DR)⁺⁺单核细胞绝对计数、IgD⁻CD27⁻B细胞占B细胞的百分比、IgD⁻CD27⁻B细胞上的CD25表达、CD8⁺T细胞占T细胞的百分比、HLA-DR⁺⁺单核细胞占单核细胞的百分比、CD14⁺CD16⁻单核细胞上的HLA-DR表达、CD14⁺单核细胞上的HLA-DR表达、IgD⁺CD38⁺B细胞上的CD20表达、CD14⁻CD16⁺单核细胞上的程序性死亡配体-1(PD-L1)表达、CD28⁺CD45RA⁻CD8dimT细胞百分比、效应记忆CD8⁺T细胞占T细胞的百分比、CD25⁺⁺CD45RA⁻CD4非调节性T细胞百分比、未成熟髓源性抑制细胞上的CD45表达与HS形成风险均呈显著负相关(OR分别为0.847、0.878、0.891、0.894、0.894、0.903、0.908、0.911、0.911、0.916、0.931、0.932、0.940、0.942,95%CI分别为0.731~0.982、0.776~0.994、0.798~0.995、0.804~0.994、0.824~0.970、0.830~0.982、0.848~0.971、0.849~0.976、0.851~0.976、0.846~0.992、0.886~0.977、0.876~0.991、0.886~0.997、0.889~0.998,P<0.05),CD14⁺CD16⁺单核细胞上的HLA-DR表达、记忆B细胞绝对计数、CD45RA⁻CD4非调节性T细胞上的CD25表达、IgD⁺CD38⁺B细胞上的CD24表达、自然杀伤细胞上的侧向散射光面积表达、CD14⁻CD16⁻细胞上的PD-L1表达、CD25⁺⁺CD4⁺T细胞占T细胞的百分比、自然杀伤细胞上的CD16⁻CD56表达、T细胞绝对计数与HS形成风险均呈显著正相关(OR分别为1.040、1.056、1.077、1.100、1.102、1.102、1.104、1.113、1.156,95%CI分别为1.001~1.080、1.001~1.114、1.020~1.138、1.030~1.174、1.008~1.205、1.024~1.187、1.016~1.200、1.034~1.198、1.047~1.276,P<0.05)。上述关联均不存在显著异质性或水平多效性(P>0.05)。厚壁菌门-梭菌纲-梭菌目-梭菌科对HS的显著总体保护效应(总效应β=-0.198,95%CI为-0.375~-0.021,P<0.05)部分通过HLA-DR⁺⁺单核细胞百分比、CD14⁺CD16⁻单核细胞上的HLA-DR表达介导(中介效应β值分别为-0.016、-0.020,95%CI分别为-0.035~-0.001、-0.050~-0.001,P值均<0.05),其中介比例分别为8.333%、13.333%。 结论 7种肠道菌群特征和23种免疫细胞表型与HS形成风险显著相关,特定肠道菌群如梭菌科、双歧杆菌目可能通过调节单核细胞HLA-DR表达等免疫细胞表型降低HS形成风险。 Abstract:Objective To investigate the causal relationships between human gut microbiota features, immune cell phenotypes, and hypertrophic scar (HS). Methods This was a study based on two-step two-sample mediation Mendelian randomization (MR) analysis. Data on human gut microbiota features, immune cell phenotypes, and HS were retrieved from the genome-wide association study database. The inverse variance weighted method was used to assess the causal relationships between 119 gut microbiota features, 731 immune cell phenotypes, and HS. Heterogeneity and horizontal pleiotropy were evaluated using Cochran's Q test and MR-Egger regression test, respectively. A two-step MR approach was employed to quantify the mediating effect of immune cell phenotypes in the association between gut microbiota features and HS. Results Seven gut microbiota features showed significant causal relationships with the risk of HS formation. Among them, the followings were significantly negatively associated with the risk of HS formation, including Actinobacteria-Actinobacteria-Bifidobacteriales, Actinobacteria-Actinobacteria-Bifidobacteriales-Bifidobacteriaceae, Bacteroidetes-Bacteroidia-Bacteroidales-Rikenellaceae-Alistipes-Alistipes senegalensis, Firmicutes-Clostridia-Clostridiales-Clostridiaceae, superpathway of heme biosynthesis from glycine, and peptidoglycan biosynthesis Ⅰ (containing meso-diaminopimelate), with OR of 0.804, 0.804, 0.784, 0.820, 0.864, and 0.686, respectively, 95% CI of 0.649-0.996, 0.649-0.996, 0.623-0.988, 0.687-0.980, 0.759-0.984, and 0.491-0.959, respectively, P<0.05. Firmicutes-Clostridia-Clostridiales-Eubacteriaceae-Eubacterium-Eubacterium eligens was significantly positively associated with the risk of HS formation (OR=1.239, with 95% CI of 1.007-1.525, P<0.05). Twenty-three immune cell phenotypes showed significant causal relationships with the risk of HS formation. These included the percentage of IgD⁻CD38⁻ B cells among B cells, absolute count of CD11c⁺ human leukocyte antigen DR (HLA-DR)⁺⁺ monocyte, percentage of IgD⁻CD27⁻ B cells among B cells, CD25 expression on IgD⁻CD27⁻ B cells, percentage of CD8⁺ T cells among T cells, percentage of HLA-DR⁺⁺ monocytes among monocytes, HLA-DR expression on CD14⁺CD16⁻ monocytes, HLA-DR expression on CD14⁺ monocytes, CD20 expression on IgD⁺CD38⁺ B cells, programmed death-ligand 1 (PD-L1) expression on CD14⁻CD16⁺ monocytes, percentage of CD28⁺CD45RA⁻CD8dim T cell, percentage of effector memory CD8⁺ T cells among T cells, percentage of CD25⁺⁺CD45RA⁻ CD4 non-regulatory T cells, and CD45 expression on immature myeloid-derived suppressor cells were significantly negatively associated with the risk of HS formation (with OR of 0.847, 0.878, 0.891, 0.894, 0.894, 0.903, 0.908, 0.911, 0.911, 0.916, 0.931, 0.932, 0.940, and 0.942, respectively, 95% CI of 0.731-0.982, 0.776-0.994, 0.798-0.995, 0.804-0.994, 0.824-0.970, 0.830-0.982, 0.848-0.971, 0.849-0.976, 0.851-0.976, 0.846-0.992, 0.886-0.977, 0.876-0.991, 0.886-0.997, and 0.889-0.998, respectively, P<0.05). HLA-DR expression on CD14⁺CD16⁺ monocytes, absolute count of memory B cells, CD25 expression on CD45RA⁻CD4 non-regulatory T cells, CD24 expression on IgD⁺CD38⁺ B cells, side scatter light area expression on natural killer (NK) cells, PD-L1 expression on CD14⁻CD16⁻ cells, percentage of CD25⁺⁺CD4⁺ T cells among T cells, CD16⁻CD56 expression on NK cells, and absolute T cell count were significantly positively associated with the risk of HS formation (with OR of 1.040, 1.056, 1.077, 1.100, 1.102, 1.102, 1.104, 1.113, and 1.156, respectively, 95% CI of 1.001-1.080, 1.001-1.114, 1.020-1.138, 1.030-1.174, 1.008-1.205, 1.024-1.187, 1.016-1.200, 1.034-1.198, and 1.047-1.276, respectively, P<0.05). No significant heterogeneity or horizontal pleiotropy was observed for the aforementioned associations (P>0.05). The significant overall protective effect of Firmicutes-Clostridia-Clostridiales-Clostridiaceae on HS (total effect β=-0.198, with 95% CI of -0.375 to -0.021, P<0.05) was partially mediated by percentage of HLA-DR⁺⁺ monocyte and HLA-DR expression on CD14⁺CD16⁻ monocytes (with mediation effect β values of -0.016 and -0.020, respectively, 95% CI of -0.035 to -0.001 and -0.050 to -0.001, respectively, P values both <0.05), with mediation proportions of 8.333% and 13.333%, respectively. Conclusions Seven gut microbiota features and 23 immune cell phenotypes are significantly associated with the risk of HS formation. Specific gut microbiota, such as Clostridiaceae and Bifidobacteriales, may reduce the risk of HS formation risk potentially by modulating immune cell phenotypes including HLA-DR expression in monocytes. -
参考文献
(43) [1] ShuchengH,LiJ,LiuYL,et al.Causal relationship between gut microbiota and pathological scars: a two-sample Mendelian randomization study[J].Front Med (Lausanne),2024,11:1405097.DOI: 10.3389/fmed.2024.1405097. [2] LiuH,LiJ,GuanC,et al.Endometriosis is a disease of immune dysfunction, which could be linked to microbiota[J].Front Genet,2024,15:1386411.DOI: 10.3389/fgene.2024.1386411. [3] ChengX,ChengB,JinR,et al.The role of circulating metabolites and gut microbiome in hypertrophic scar: a two-sample Mendelian randomization study[J].Arch Dermatol Res,2024,316(6):315.DOI: 10.1007/s00403-024-03116-8. [4] ClaytonSM,ShafikhaniSH,SoulikaAM.Macrophage and neutrophil dysfunction in diabetic wounds[J].Adv Wound Care (New Rochelle),2024,13(9):463-484.DOI: 10.1089/wound.2023.0149. [5] LivantsovaEN,LeonovGE,StarodubovaAV,et al.Diet and the gut microbiome as determinants modulating metabolic outcomes in young obese adults[J].Biomedicines,2024,12(7):1601.DOI: 10.3390/biomedicines12071601. [6] LiuX,LiuH,WuX,et al.Xiaoyaosan against depression through suppressing LPS mediated TLR4/NLRP3 signaling pathway in "microbiota-gut-brain" axis[J].J Ethnopharmacol,2024,335:118683.DOI: 10.1016/j.jep.2024.118683. [7] Réthi-NagyZ,JuhászS.Microbiome's universe: impact on health, disease and cancer treatment[J].J Biotechnol,2024,392:161-179.DOI: 10.1016/j.jbiotec.2024.07.002. [8] Herrera-LuisE,OrtegaVE,AmplefordEJ,et al.Multi-ancestry genome-wide association study of asthma exacerbations[J].Pediatr Allergy Immunol,2022,33(6):e13802.DOI: 10.1111/pai.13802. [9] GuoW,ZhaoL,HuangW,et al.Sodium-glucose cotransporter 2 inhibitors, inflammation, and heart failure: a two-sample Mendelian randomization study[J].Cardiovasc Diabetol,2024,23(1):118.DOI: 10.1186/s12933-024-02210-5. [10] Lopera-MayaEA,KurilshikovA,van der GraafA,et al.Effect of host genetics on the gut microbiome in 7,738 participants of the Dutch Microbiome Project[J].Nat Genet,2022,54(2):143-151.DOI: 10.1038/s41588-021-00992-y. [11] KurkiMI,KarjalainenJ,PaltaP,et al.FinnGen provides genetic insights from a well-phenotyped isolated population[J].Nature,2023,613(7944):508-518.DOI: 10.1038/s41586-022-05473-8. [12] OrrùV,SteriM,SidoreC,et al.Complex genetic signatures in immune cells underlie autoimmunity and inform therapy[J].Nat Genet,2020,52(10):1036-1045.DOI: 10.1038/s41588-020-0684-4. [13] LouJ,CuiS,LiJ,et al.Causal relationship between the gut microbiome and basal cell carcinoma, melanoma skin cancer, ease of skin tanning: evidence from three two-sample mendelian randomisation studies[J].Front Immunol,2024,15:1279680.DOI: 10.3389/fimmu.2024.1279680. [14] BurgessS,ThompsonSG, Genetics CollaborationCRP CHD. Methods for meta-analysis of individual participant data from Mendelian randomisation studies with binary outcomes[J].Stat Methods Med Res,2016,25(1):272-293.DOI: 10.1177/0962280212451882. [15] BurgessS,WoolfB,MasonAM,et al.Addressing the credibility crisis in Mendelian randomization[J].BMC Med,2024,22(1):374.DOI: 10.1186/s12916-024-03607-5. [16] WoolfB,GillD,GrantAJ,et al.MVMRmode: introducing an R package for plurality valid estimators for multivariable Mendelian randomisation[J].PLoS One,2024,19(5):e0291183.DOI: 10.1371/journal.pone.0291183. [17] YuanS,LarssonSC,GillD,et al.Concerns about instrumental variable selection for biological effect versus uptake of proton pump inhibitors in Mendelian randomisation analysis[J].Gut,2024,74(1):e6.DOI: 10.1136/gutjnl-2024-332280. [18] ShenHT,FangYT,TsaiWH,et al.A Lactobacillus combination ameliorates lung inflammation in an elastase/LPS-induced mouse model of chronic obstructive pulmonary disease[J].Probiotics Antimicrob Proteins,2025,17(5):3416-3428.DOI: 10.1007/s12602-024-10300-9. [19] JungY,ParkC,LeeH,et al.Association of the skin microbiome with the biomechanical scar properties in patients with burns[J].Burns,2025,51(3):107372.DOI: 10.1016/j.burns.2025.107372. [20] 陈文涛,王小祥,郑文炼,等.基于双样本孟德尔随机化分析探索人肠道菌群与增生性瘢痕之间的因果关系[J].中华烧伤与创面修复杂志,2024,40(4):333-341.DOI: 10.3760/cma.j.cn501225-20231129-00215. [21] CadyN,PetersonSR,FreedmanSN,et al.Beyond metabolism: the complex interplay between dietary phytoestrogens, gut bacteria, and cells of nervous and immune systems[J].Front Neurol,2020,11:150.DOI: 10.3389/fneur.2020.00150. [22] ZengY,FanN,GuX,et al.Characteristics of gut microbiota and serum metabolism in patients with atopic dermatitis[J].Skin Res Technol,2024,30(7):e13792.DOI: 10.1111/srt.13792. [23] YangL,LiD,SunS,et al.Dupilumab therapy improves gut microbiome dysbiosis and tryptophan metabolism in Chinese patients with atopic dermatitis[J].Int Immunopharmacol,2024,131:111867.DOI: 10.1016/j.intimp.2024.111867. [24] YegorovS,BabenkoD,KozhakhmetovS,et al.Psoriasis is associated with elevated gut IL-1α and intestinal microbiome alterations[J].Front Immunol,2020,11:571319.DOI: 10.3389/fimmu.2020.571319. [25] ClimentE,Martinez-BlanchJF,LlobregatL,et al.Changes in gut microbiota correlates with response to treatment with probiotics in patients with atopic dermatitis. A post hoc analysis of a clinical trial[J].Microorganisms,2021,9(4):854.DOI: 10.3390/microorganisms9040854. [26] StefiaLV,LeeJ,PatelJ,et al.Secretome components from Faecalibacterium prausnitzii strains A2-165 and AHMP21 modulate cutaneous wound inflammation[J].J Invest Dermatol,2020,140(11):2312-2315.e6.DOI: 10.1016/j.jid.2020.02.027. [27] TsakmaklisA,FarowskiF,ZennerR,et al.TIGIT+ NK cells in combination with specific gut microbiota features predict response to checkpoint inhibitor therapy in melanoma patients[J].BMC Cancer,2023,23(1):1160.DOI: 10.1186/s12885-023-11551-5. [28] XueK,ZhangG,LiZ,et al.Dissecting the association between gut microbiota and hypertrophic scarring: a bidirectional Mendelian randomization study[J].Front Microbiol,2024,15:1345717.DOI: 10.3389/fmicb.2024.1345717. [29] MunleyJA,KellyLS,ParkG,et al.Acute emergence of the intestinal pathobiome after postinjury pneumonia[J].J Trauma Acute Care Surg,2024,97(1):65-72.DOI: 10.1097/TA.0000000000004300. [30] ShangguanJ,YuF,DingB,et al.Hydrogel-forming viscous liquid in response to ROS restores the gut mucosal barrier of colitis mice via regulating oxidative redox homeostasis[J].Acta Biomater,2024,184:127-143.DOI: 10.1016/j.actbio.2024.06.021. [31] ShearerJ,ShahS,Shen-TuG,et al.Microbial features linked to medication strategies in cardiometabolic disease management[J].ACS Pharmacol Transl Sci,2024,7(4):991-1001.DOI: 10.1021/acsptsci.3c00261. [32] LiuS,ZhaoW,LiuX,et al.Metagenomic analysis of the gut microbiome in atherosclerosis patients identify cross-cohort microbial signatures and potential therapeutic target[J].FASEB J,2020,34(11):14166-14181.DOI: 10.1096/fj.202000622R. [33] ZhangY,ChengS,ZouH,et al.Correlation of the gut microbiome and immune-related adverse events in gastrointestinal cancer patients treated with immune checkpoint inhibitors[J].Front Cell Infect Microbiol,2023,13:1099063.DOI: 10.3389/fcimb.2023.1099063. [34] WangZ,GuoM,LiJ,et al.Composition and functional profiles of gut microbiota reflect the treatment stage, severity, and etiology of acute pancreatitis[J].Microbiol Spectr,2023,11(5):e0082923.DOI: 10.1128/spectrum.00829-23. [35] HiguchiBS,RodriguesN,GonzagaMI,et al.Intestinal dysbiosis in autoimmune diabetes is correlated with poor glycemic control and increased interleukin-6: a pilot study[J].Front Immunol,2018,9:1689.DOI: 10.3389/fimmu.2018.01689. [36] ZhaoR,LiY,HanM,et al.Genetic causality linking skin microbiota to skin cancer: Mendelian randomization study and meta-analysis[J].Medicine (Baltimore),2025,104(32):e43571.DOI: 10.1097/MD.0000000000043571. [37] LoffredoLF,SavageTM,RinghamOR,et al.Treg-tissue cell interactions in repair and regeneration[J].J Exp Med,2024,221(6):e20231244.DOI: 10.1084/jem.20231244. [38] TredgetEE,YangL,DelehantyM,et al.Polarized Th2 cytokine production in patients with hypertrophic scar following thermal injury[J].J Interferon Cytokine Res,2006,26(3):179-189.DOI: 10.1089/jir.2006.26.179. [39] IslamN,WhitehouseM,MehendaleS,et al.Post-traumatic immunosuppression is reversed by anti-coagulated salvaged blood transfusion: deductions from studying immune status after knee arthroplasty[J].Clin Exp Immunol,2014,177(2):509-520.DOI: 10.1111/cei.12351. [40] KimD,LoE,KimD,et al.Regulatory T cells conditioned media stimulates migration in HaCaT keratinocytes: involvement of wound healing[J].Clin Cosmet Investig Dermatol,2020,13:443-453.DOI: 10.2147/CCID.S252778. [41] ChancheewaB,AsawanondaP,NoppakunN,et al.Myofibroblasts, B cells, and mast cells in different types of long-standing acne scars[J].Skin Appendage Disord,2022,8(6):469-475.DOI: 10.1159/000524566. [42] 甘文军,王婧薷,何佳,等.人免疫细胞表型与瘢痕疙瘩之间因果关系的双样本孟德尔随机化分析[J].中华烧伤与创面修复杂志,2025,41(1):84-93.DOI: 10.3760/cma.j.cn501225-20231130-00219. [43] 李涛,朱晨晨,陈今源,等.人炎症蛋白与瘢痕疙瘩之间因果关系的孟德尔随机化分析[J].中华烧伤与创面修复杂志,2025,41(2):180-187.DOI: 10.3760/cma.j.cn501225-20240526-00198. -

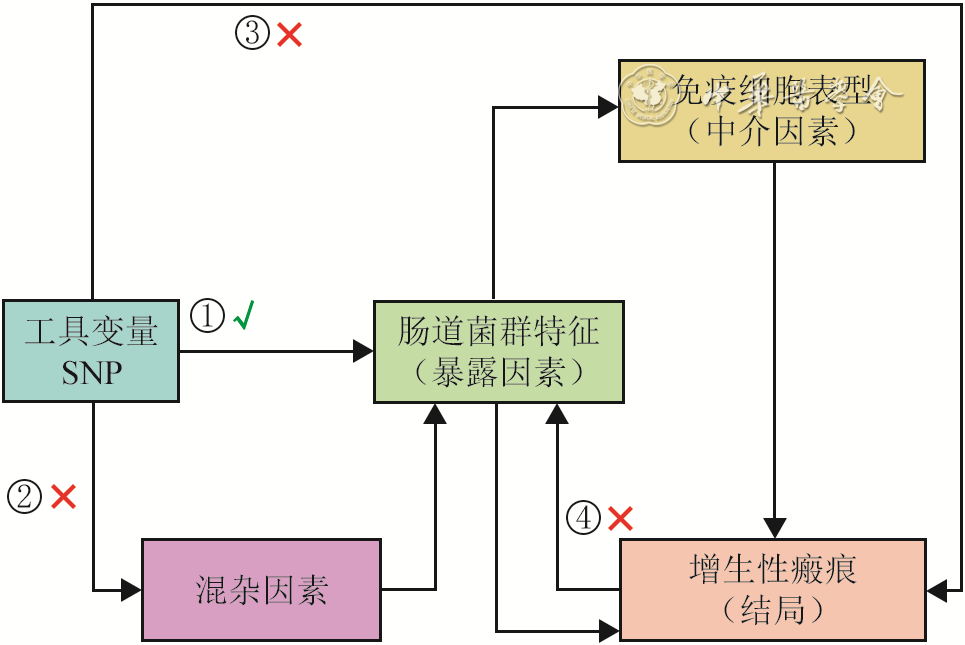
图 1 人肠道菌群特征和免疫细胞表型与增生性瘢痕之间因果关系的两步双样本中介孟德尔随机化分析的核心假设和分析流程图
注:SNP为单核苷酸多态性;“√”表示存在关联或有效路径,“×”表示该路径不存在或已被排除;①表示关联性假设,即工具变量与暴露因素强相关;②表示独立性假设,即工具变量与混杂因素无关;③表示排他性假设,即工具变量不通过暴露因素以外的途径影响结局;④表示无反向因果,即结局不会反向影响暴露因素
Table 1. 逆方差加权法分析得出7种人肠道菌群特征与增生性瘢痕存在显著因果关系
肠道菌群特征 SNP数(个) OR 95%CI P值 放线菌门-放线菌纲-双歧杆菌目 13 0.804 0.649~0.996 0.046 放线菌门-放线菌纲-双歧杆菌目-双歧杆菌科 13 0.804 0.649~0.996 0.046 拟杆菌门-拟杆菌纲-拟杆菌目-理研菌科-阿尔斯提普斯属-塞内加尔阿尔斯提普斯种 12 0.784 0.623~0.988 0.039 厚壁菌门-梭菌纲-梭菌目-梭菌科 10 0.820 0.687~0.980 0.029 甘氨酸起始的血红素生物合成超途径 10 0.864 0.759~0.984 0.028 含内消旋二氨基庚二酸的肽聚糖生物合成Ⅰ 10 0.686 0.491~0.959 0.028 厚壁菌门-梭菌纲-梭菌目-真杆菌科-真杆菌属-纤毛真杆菌种 14 1.239 1.007~1.525 0.043 注:双歧杆菌目为单型目,仅包含双歧杆菌科,故两者遗传信号完全重叠,结果一致;SNP为单核苷酸多态性  下载: 导出CSV
下载: 导出CSV
Table 2. 逆方差加权法分析得出23种人免疫细胞表型与增生性瘢痕存在显著因果关系
免疫细胞表型 SNP数(个) OR 95%CI P值 IgD⁻CD38⁻B细胞占B细胞的百分比 22 0.847 0.731~0.982 0.028 CD11c⁺HLA-DR⁺⁺单核细胞绝对计数 18 0.878 0.776~0.994 0.040 IgD⁻CD27⁻B细胞占B细胞的百分比 25 0.891 0.798~0.995 0.041 IgD⁻CD27⁻B细胞上的CD25表达 20 0.894 0.804~0.994 0.039 CD8⁺T细胞占T细胞的百分比 28 0.894 0.824~0.970 0.007 HLA-DR⁺⁺单核细胞占单核细胞的百分比 15 0.903 0.830~0.982 0.017 CD14⁺CD16⁻单核细胞上的HLA-DR表达 19 0.908 0.848~0.971 0.005 CD14⁺单核细胞上的HLA-DR表达 17 0.911 0.849~0.976 0.008 IgD⁺CD38⁺B细胞上的CD20表达 24 0.911 0.851~0.976 0.008 CD14⁻CD16⁺单核细胞上的PD-L1表达 21 0.916 0.846~0.992 0.030 CD28⁺CD45RA⁻CD8dimT细胞百分比 16 0.931 0.886~0.977 0.004 效应记忆CD8⁺T细胞占T细胞的百分比 23 0.932 0.876~0.991 0.025 CD25⁺⁺CD45RA⁻CD4非调节性T细胞百分比 26 0.940 0.886~0.997 0.040 未成熟髓源性抑制细胞上的CD45表达 14 0.942 0.889~0.998 0.044 CD14⁺CD16⁺单核细胞上的HLA-DR表达 18 1.040 1.001~1.080 0.044 记忆B细胞绝对计数 22 1.056 1.001~1.114 0.044 CD45RA⁻CD4非调节性T细胞上的CD25表达 19 1.077 1.020~1.138 0.007 IgD⁺CD38⁺B细胞上的CD24表达 25 1.100 1.030~1.174 0.004 自然杀伤细胞上的侧向散射光面积表达 12 1.102 1.008~1.205 0.033 CD14⁻CD16⁻细胞上的PD-L1表达 20 1.102 1.024~1.187 0.010 CD25⁺⁺CD4⁺T细胞占T细胞的百分比 16 1.104 1.016~1.200 0.020 自然杀伤细胞上的CD16⁻CD56表达 13 1.113 1.034~1.198 0.004 T细胞绝对计数 30 1.156 1.047~1.276 0.004 注:HLA-DR为人类白细胞抗原-DR,PD-L1为程序性死亡配体-1,SNP为单核苷酸多态性
下载: 导出CSV
Table 3. 逆方差加权法分析得出8组具有显著因果关系的人肠道菌群特征-免疫细胞表型对
肠道菌群特征 SNP数(个) 免疫细胞表型 OR 95%CI P值 甘氨酸起始的血红素生物合成超途径 10 CD14⁻CD16⁻细胞上的PD-L1表达 1.146 1.050~1.251 0.002 厚壁菌门-梭菌纲-梭菌目-梭菌科 12 HLA-DR⁺⁺单核细胞百分比 1.202 1.051~1.375 0.007 厚壁菌门-梭菌纲-梭菌目-梭菌科 11 CD14⁺CD16⁻单核细胞上的HLA-DR表达 1.186 1.036~1.357 0.013 厚壁菌门-梭菌纲-梭菌目-梭菌科 13 CD14⁺单核细胞上的HLA-DR表达 1.181 1.031~1.352 0.016 厚壁菌门-梭菌纲-梭菌目-梭菌科 14 CD39⁺CD4⁺调节性T细胞上的CD3表达 1.363 1.088~1.708 0.007 厚壁菌门-梭菌纲-梭菌目-梭菌科 12 髓系树突状细胞上的CD80表达 1.139 1.008~1.287 0.037 拟杆菌门-拟杆菌纲-拟杆菌目-理研菌科-阿尔斯提普斯属-塞内加尔阿尔斯提普斯种 9 CD45RA⁺CD8⁺T细胞绝对计数 0.816 0.693~0.960 0.014 拟杆菌门-拟杆菌纲-拟杆菌目-理研菌科-阿尔斯提普斯属-塞内加尔阿尔斯提普斯种 8 CD28⁺CD45RA⁻CD8dimT细胞百分比 1.233 1.001~1.518 0.049 注:SNP为单核苷酸多态性;PD-L1为程序性死亡配体-1,HLA-DR为人类白细胞抗原-DR
下载: 导出CSV
-

 下载:
下载:


图(2) / 表(3)
计量
- 文章访问数: 571
- HTML全文浏览量: 341
- PDF下载量: 17
- 被引次数: 0
