




Effect and mechanism of astaxanthin on the aging of high glucose-treated human skin fibroblasts
-
摘要:
目的 探讨虾青素对高糖处理的人皮肤成纤维细胞(Fb)衰老的作用及其机制。 方法 该研究为实验研究。取人皮肤Fb,分为常规培养的对照组、用终物质的量浓度30 mmol/L的葡萄糖处理的高糖组及分别用终物质的量浓度25、50 μmol/L的虾青素预处理后再同高糖组处理的低虾青素组、高虾青素组。培养48 h后,采用细胞计数试剂盒-8检测细胞存活率,用荧光探针法检测细胞中活性氧水平,采用比色法检测细胞中丙二醛、谷胱甘肽、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、谷胱甘肽过氧化物酶(GSH-Px)水平,采用蛋白质印迹法检测细胞中p53、磷酸化p53、p21、p16、Rb、磷酸化Rb、基质金属蛋白酶1(MMP1)、MMP3、MMP13蛋白表达水平及细胞质和细胞核中核转录因子红系2相关因子2(Nrf2)蛋白表达水平。另取人皮肤Fb,分为同前处理的对照组、高糖组、高虾青素组及用终物质的量浓度为50 μmol/L的虾青素和10 μmol/L的ML385预处理后再同高糖组细胞处理的高虾青素+ML385组。培养48 h后,同前检测细胞存活率,细胞中p53、磷酸化p53、p21、p16、Rb、磷酸化Rb、MMP1、MMP3、MMP13蛋白表达水平及细胞质和细胞核中Nrf2蛋白表达水平。以上实验中样本数均为3。 结果 培养48 h后,与对照组比较,高糖组细胞存活率明显降低(P<0.05);与高糖组比较,低虾青素组和高虾青素组细胞存活率均明显升高(P<0.05);与低虾青素组比较,高虾青素组细胞存活率明显升高(P<0.05)。与对照组比较,高糖组细胞中活性氧、丙二醛水平均明显升高(P<0.05),谷胱甘肽、SOD、CAT、GSH-Px水平均明显降低(P<0.05)。与高糖组比较,低虾青素组细胞中活性氧、丙二醛水平均明显降低(P<0.05),CAT、GSH-Px水平均明显升高(P<0.05);高虾青素组细胞中活性氧、丙二醛水平均明显降低(P<0.05),谷胱甘肽、SOD、CAT、GSH-Px水平均明显升高(P<0.05)。与低虾青素组比较,高虾青素组细胞中活性氧、丙二醛水平均明显降低(P<0.05),谷胱甘肽、CAT、GSH-Px水平均明显升高(P<0.05)。与对照组比较,高糖组细胞中p53、p21、磷酸化p53、p16、Rb、MMP1、MMP3、MMP13蛋白表达水平均明显升高(P<0.05),磷酸化Rb蛋白表达水平明显降低(P<0.05)。与高糖组比较,低虾青素组细胞中p53、p21、磷酸化p53、MMP13蛋白表达水平均明显降低(P<0.05),磷酸化Rb蛋白表达水平明显升高(P<0.05);高虾青素组细胞中p53、p21、磷酸化p53、p16、Rb、MMP1、MMP3、MMP13蛋白表达水平均明显降低(P<0.05),磷酸化Rb蛋白表达水平明显升高(P<0.05)。与低虾青素组比较,高虾青素组细胞中p21、磷酸化p53、p16、Rb、MMP1蛋白表达水平均明显降低(P<0.05)。与对照组、高虾青素组比较,高糖组细胞质中Nrf2蛋白表达水平明显升高(P值均<0.05),细胞核中Nrf2蛋白表达水平明显降低(P值均<0.05)。与低虾青素组比较,高糖组细胞质中Nrf2蛋白表达水平明显升高(P<0.05);高虾青素组细胞质中Nrf2蛋白表达水平明显降低(P<0.05),细胞核中Nrf2蛋白表达水平明显升高(P<0.05)。培养48 h后,与对照组[(100.0±6.0)%]比较,高糖组细胞存活率[(73.9±2.2)%]明显降低(P<0.05);与高糖组比较,高虾青素组细胞存活率[(93.8±1.5)%]明显升高(P<0.05);与高虾青素组比较,高虾青素+ML385组细胞存活率[(71.7±2.7)%]明显降低(P值均<0.05)。与对照组、高虾青素组比较,高糖组细胞中p53、磷酸化p53、p21、p16、Rb、MMP1、MMP3、MMP13蛋白表达水平均明显升高(P<0.05),磷酸化Rb蛋白表达水平明显降低(P值均<0.05);与高虾青素组比较,高虾青素+ML385组细胞中p53、磷酸化p53、p21、p16、Rb、MMP1、MMP13蛋白表达水平均明显升高(P<0.05),磷酸化Rb蛋白表达水平明显降低(P<0.05)。与对照组比较,高糖组细胞质中Nrf2蛋白表达水平明显升高(P<0.05);与高糖组比较,高虾青素组细胞质中Nrf2蛋白表达水平明显降低(P<0.05),细胞核中Nrf2蛋白表达水平明显升高(P<0.05);与高虾青素组比较,高虾青素+ML385组细胞核中Nrf2蛋白表达水平明显降低(P<0.05)。 结论 虾青素通过调节Nrf2核位移抑制高糖处理的人皮肤Fb氧化应激反应,下调细胞衰老相关蛋白的表达,从而缓解细胞衰老。 Abstract:Objective To investigate the effect and mechanism of astaxanthin on the aging of high glucose-treated human skin fibroblasts (Fbs). Methods The study was an experimental study. Human skin Fbs were collected and divided into control group with conventional culture, high glucose group treated with glucose at a final molarity of 30 mmol/L, and low astaxanthin group and high astaxanthin group pretreated with astaxanthin at final molarities of 25 and 50 μmol/L respectively and then treated as that in high glucose group. After 48 h of culture, the cell survival rate was detected by the cell counting kit-8, the reactive oxygen species level in cells was detected by the fluorescent probe method, the levels of malondialdehyde, glutathione, superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GSH-Px) in cells were detected by colorimetry, and the protein expression levels of p53, phosphorylated p53, p21, p16, Rb, phosphorylated Rb, matrix metalloproteinase 1 (MMP1), MMP3, and MMP13 in cells and nuclear factor-erythroid 2-related factor 2 (Nrf2) in cytoplasm and nucleus were detected by Western blotting. Another batch of human skin Fbs were collected and divided into control group, high glucose group, and high astaxanthin group which were treated as before, and high astaxanthin+ML385 group, which were pretreated with astaxanthin at a final molarity of 50 μmol/L and ML385 at a final molarity of 10 μmol/L respectively and then treated as that in high glucose group. After 48 h of culture, the cell survival rate, the protein expression levels of p53, phosphorylated p53, p21, p16, Rb, phosphorylated Rb, MMP1, MMP3, and MMP13 in cells and Nrf2 in cytoplasm and nucleus were detected as before. The sample number in all experiments above was 3. Results After 48 h of culture, compared with that in control group, the cell survival rate in high glucose group was significantly decreased (P<0.05); compared with that in high glucose group, the cell survival rates in low astaxanthin group and high astaxanthin group were significantly increased (P<0.05); compared with that in low astaxanthin group, the cell survival rate in high astaxanthin group was significantly increased (P<0.05). Compared with those in control group, the levels of reactive oxygen species and malondialdehyde in cells were significantly increased (P<0.05), while the levels of glutathione, SOD, CAT, and GSH-Px in cells were significantly decreased in high glucose group (P<0.05). Compared with those in high glucose group, the levels of reactive oxygen species and malondialdehyde in cells were significantly decreased (P<0.05), while the levels of CAT and GSH-Px in cells were significantly increased in low astaxanthin group (P<0.05); the levels of reactive oxygen species and malondialdehyde in cells were significantly decreased (P<0.05), while the levels of glutathione, SOD, CAT, and GSH-Px in cells were significantly increased in high astaxanthin group (P<0.05). Compared with those in low astaxanthin group, the levels of reactive oxygen species and malondialdehyde in cells were significantly decreased (P<0.05), while the levels of glutathione, CAT, and GSH-Px in cells were significantly increased in high astaxanthin group (P<0.05). Compared with those in control group, the protein expression levels of p53, p21, phosphorylated p53, p16, Rb, MMP1, MMP3, and MMP13 in cells were significantly increased (P<0.05), while the protein expression level of phosphorylated Rb in cells was significantly decreased in high glucose group (P<0.05). Compared with those in high glucose group, the protein expression levels of p53, p21, phosphorylated p53, and MMP13 in cells were significantly decreased (P<0.05), while the protein expression level of phosphorylated Rb in cells was significantly increased in low astaxanthin group (P<0.05); the protein expression levels of p53, p21, phosphorylated p53, p16, Rb, MMP1, MMP3, and MMP13 in cells were significantly decreased (P<0.05), while the protein expression level of phosphorylated Rb in cells was significantly increased in high astaxanthin group (P<0.05). Compared with those in low astaxanthin group, the protein expression levels of p21, phosphorylated p53, p16, Rb, and MMP1 in cells in high astaxanthin group were significantly decreased (P<0.05). Compared with those in control group and high astaxanthin group, the protein expression level of cytoplasmic Nrf2 was significantly increased (with P values both <0.05), while the protein expression level of nuclear Nrf2 was significantly decreased in high glucose group (with P values both <0.05). Compared with those in low astaxanthin group, the protein expression level of cytoplasmic Nrf2 was significantly increased in high glucose group (P<0.05); the protein expression level of cytoplasmic Nrf2 was significantly decreased (P<0.05), while the protein expression level of nuclear Nrf2 was significantly increased in high astaxanthin group (P<0.05). After 48 h of culture, compared with (100.0±6.0)% in control group, the cell survival rate was significantly decreased in high glucose group ((73.9±2.2)%, P<0.05); compared with that in high glucose group, the cell survival rate was significantly increased in high astaxanthin group ((93.8±1.5)%, P<0.05); compared with that in high astaxanthin group, the cell survival rate was significantly decreased in high astaxanthin+ML385 group ((71.7±2.7)%, P<0.05). Compared with those in control group and high astaxanthin group, the protein expression levels of p53, phosphorylated p53, p21, p16, Rb, MMP1, MMP3, and MMP13 in cells were significantly increased (P<0.05), while the protein expression level of phosphorylated Rb in cells was significantly decreased in high glucose group (with P values both <0.05); compared with those in high astaxanthin group, the protein expression levels of p53, phosphorylated p53, p21, p16, Rb, MMP1, and MMP13 in cells were significantly increased (P<0.05), while the protein expression level of phosphorylated Rb in cells was significantly decreased in high astaxanthin+ML385 group (P<0.05). Compared with that in control group, the protein expression level of cytoplasmic Nrf2 was significantly increased in high glucose group (P<0.05); compared with those in high glucose group, the protein expression level of cytoplasmic Nrf2 was significantly decreased (P<0.05), while the protein expression level of nuclear Nrf2 was significantly increased in high astaxanthin group (P<0.05); compared with that in high astaxanthin group, the protein expression level of nuclear Nrf2 was significantly decreased in high astaxanthin+ML385 group (P<0.05). Conclusions Astaxanthin can alleviate aging of high glucose-treated human skin Fbs by regulating Nrf2 nuclear translocation to inhibit oxidative stress and downregulate the expression of aging-related proteins. -
参考文献
(40) [1] TomicD, ShawJE, MaglianoDJ. The burden and risks of emerging complications of diabetes mellitus[J]. Nat Rev Endocrinol, 2022, 18(9): 525-539.DOI: 10.1038/s41574-022-00690-7. [2] 李威杰,秦晓光,朱甜,等. 减重与代谢外科新进展[J]. 中华消化外科杂志,2023,22(8):958-964.DOI: 10.3760/cma.j.cn115610-20230711-00399. [3] BurgessJL, WyantWA, Abdo AbujamraB, et al. Diabetic wound-healing science[J]. Medicina (Kaunas), 2021, 57(10):1072.DOI: 10.3390/medicina57101072. [4] WolfSJ, MelvinWJ, GallagherK. Macrophage-mediated inflammation in diabetic wound repair[J]. Semin Cell Dev Biol, 2021, 119: 111-118.DOI: 10.1016/j.semcdb.2021.06.013. [5] StoneA, DonohueCM. Diabetic foot ulcers in geriatric patients[J]. Clin Geriatr Med, 2024, 40(3): 437-447.DOI: 10.1016/j.cger.2024.03.002. [6] WilkinsonHN, HardmanMJ. Wound healing: cellular mechanisms and pathological outcomes[J]. Open Biol, 2020, 10(9): 200223.DOI: 10.1098/rsob.200223. [7] DengL, DuC, SongP, et al. The role of oxidative stress and antioxidants in diabetic wound healing[J]. Oxid Med Cell Longev, 2021, 2021: 8852759.DOI: 10.1155/2021/8852759. [8] MuX, WuX, HeW, et al. Pyroptosis and inflammasomes in diabetic wound healing[J]. Front Endocrinol (Lausanne), 2022, 13: 950798.DOI: 10.3389/fendo.2022.950798. [9] NeaguM, ConstantinC, SurcelM, et al. Diabetic neuropathy: a NRF2 disease?[J]. J Diabetes, 2024, 16(9): e13524.DOI: 10.1111/1753-0407.13524. [10] ZhangDD. Thirty years of NRF2: advances and therapeutic challenges[J]. Nat Rev Drug Discov, 2025, 24(6): 421-444.DOI: 10.1038/s41573-025-01145-0. [11] 郭佳,张军霞. 核转录因子红系2相关因子2在创面愈合中的作用研究进展[J]. 中华烧伤与创面修复杂志,2023,39(1): 91-95.DOI: 10.3760/cma.j.cn501225-20220531-00209. [12] UrakazeM, KobashiC, SatouY, et al. The beneficial effects of astaxanthin on glucose metabolism and modified low-density lipoprotein in healthy volunteers and subjects with prediabetes[J]. Nutrients, 2021, 13(12):4381.DOI: 10.3390/nu13124381. [13] NishidaY, BergPC, ShakersainB, et al. Astaxanthin: past, present, and future[J]. Mar Drugs, 2023, 21(10):514.DOI: 10.3390/md21100514. [14] ZhangQ, LuoC, LiZ, et al. Astaxanthin activates the Nrf2/Keap1/HO-1 pathway to inhibit oxidative stress and ferroptosis, reducing triphenyl phosphate (TPhP)-induced neurodevelopmental toxicity[J]. Ecotoxicol Environ Saf, 2024, 271: 115960.DOI: 10.1016/j.ecoenv.2024.115960. [15] HafezMH, El-FarAH, ElblehiSS. Astaxanthin alleviates fipronil-induced neuronal damages in male rats through modulating oxidative stress, apoptosis, and inflammatory markers[J]. Sci Rep, 2025, 15(1): 14299.DOI: 10.1038/s41598-025-95447-3. [16] ZhuY, RuanCX, WangJ, et al. High glucose inhibits the survival of HRMCs and its mechanism[J]. Eur Rev Med Pharmacol Sci, 2022, 26(16): 5683-5688.DOI: 10.26355/eurrev_202208_29502. [17] ZhangZ, QiuY, LiW, et al. Astaxanthin alleviates foam cell formation and promotes cholesterol efflux in ox-LDL-induced RAW264.7 cells via circTPP2/miR-3073b-5p/ABCA1 pathway[J]. Molecules, 2023, 28(4):1701.DOI: 10.3390/molecules28041701. [18] YangCS, GuoXS, YueYY, et al. Astaxanthin promotes the survival of adipose-derived stem cells by alleviating oxidative stress via activating the Nrf2 signaling pathway[J]. Int J Mol Sci, 2023, 24(4):3850.DOI: 10.3390/ijms24043850. [19] JuCC, LiuXX, LiuLH, et al. Epigenetic modification: a novel insight into diabetic wound healing[J]. Heliyon, 2024, 10(6): e28086.DOI: 10.1016/j.heliyon.2024.e28086. [20] WangG, YangF, ZhouW, et al. The initiation of oxidative stress and therapeutic strategies in wound healing[J]. Biomed Pharmacother, 2023, 157: 114004.DOI: 10.1016/j.biopha.2022.114004. [21] LiuY, LiuY, HeW, et al. Fibroblasts: immunomodulatory factors in refractory diabetic wound healing[J]. Front Immunol, 2022, 13: 918223.DOI: 10.3389/fimmu.2022.918223. [22] FengJ, WangJ, WangY, et al. Oxidative stress and lipid peroxidation: prospective associations between ferroptosis and delayed wound healing in diabetic ulcers[J]. Front Cell Dev Biol, 2022, 10: 898657.DOI: 10.3389/fcell.2022.898657. [23] HuangK, MiB, XiongY, et al. Angiogenesis during diabetic wound repair: from mechanism to therapy opportunity[J/OL]. Burns Trauma, 2025, 13: tkae052[2025-08-18].https://pubmed.ncbi.nlm.nih.gov/39927093/.DOI: 10.1093/burnst/tkae052. [24] CitrinKM, ChaubeB, Fernández-HernandoC, et al. Intracellular endothelial cell metabolism in vascular function and dysfunction[J]. Trends Endocrinol Metab, 2025, 36(8): 744-755.DOI: 10.1016/j.tem.2024.11.004. [25] GasekNS, YanP, ZhuJ, et al. Clearance of p21 highly expressing senescent cells accelerates cutaneous wound healing[J]. Nat Aging, 2025, 5(1): 21-27.DOI: 10.1038/s43587-024-00755-4. [26] ZhangS, MengN, LiuS, et al. Targeting senescent HDF with the USP7 inhibitor P5091 to enhance DFU wound healing through the p53 pathway[J]. Biochem Biophys Res Commun, 2024, 722: 150149.DOI: 10.1016/j.bbrc.2024.150149. [27] SamarawickramaPN, ZhangG, ZhuE, et al. Clearance of senescent cells enhances skin wound healing in type 2 diabetic mice[J]. Theranostics, 2024, 14(14): 5429-5442.DOI: 10.7150/thno.100991. [28] McElhinneyK, IrnatenM, O'BrienC. P53 and myofibroblast apoptosis in organ fibrosis[J]. Int J Mol Sci, 2023, 24(7):6737.DOI: 10.3390/ijms24076737. [29] MaZ, DingY, DingX, et al. PDK4 rescues high-glucose-induced senescent fibroblasts and promotes diabetic wound healing through enhancing glycolysis and regulating YAP and JNK pathway[J]. Cell Death Discov, 2023, 9(1): 424.DOI: 10.1038/s41420-023-01725-2. [30] QinY, WuK, ZhangZ, et al. NLRC3 deficiency promotes cutaneous wound healing due to the inhibition of p53 signaling[J]. Biochim Biophys Acta Mol Basis Dis, 2022, 1868(11): 166518.DOI: 10.1016/j.bbadis.2022.166518. [31] JiangG, JiangT, ChenJ, et al. Mitochondrial dysfunction and oxidative stress in diabetic wound[J]. J Biochem Mol Toxicol, 2023, 37(7): e23407.DOI: 10.1002/jbt.23407. [32] QinY, LiuH, WuH. Cellular senescence in health, disease, and lens aging[J]. Pharmaceuticals (Basel), 2025, 18(2):244.DOI: 10.3390/ph18020244. [33] ChoiH, KangC. Living beyond restriction: LBR promotes cellular immortalization by suppressing genomic instability and senescence[J]. FEBS J, 2024, 291(10): 2091-2093.DOI: 10.1111/febs.17141. [34] Mahiny-ShahmohammadyD, HauckL, BilliaF. Defining the molecular underpinnings controlling cardiomyocyte proliferation[J]. Clin Sci (Lond), 2022, 136(12): 911-934.DOI: 10.1042/CS20211180. [35] O'ReillyS, MarkiewiczE, IdowuOC. Aging, senescence, and cutaneous wound healing-a complex relationship[J]. Front Immunol, 2024, 15: 1429716.DOI: 10.3389/fimmu.2024.1429716. [36] ÅgrenMS, LitmanT, EriksenJO, et al. Gene expression linked to reepithelialization of human skin wounds[J]. Int J Mol Sci, 2022, 23(24):15746.DOI: 10.3390/ijms232415746. [37] JacquierEF, KassisA, MarcuD, et al. Phytonutrients in the promotion of healthspan: a new perspective[J]. Front Nutr, 2024, 11: 1409339.DOI: 10.3389/fnut.2024.1409339. [38] KanwuguON, GlukharevaTV, DanilovaIG, et al. Natural antioxidants in diabetes treatment and management: prospects of astaxanthin[J]. Crit Rev Food Sci Nutr, 2022, 62(18): 5005-5028.DOI: 10.1080/10408398.2021.1881434. [39] 商冠华,田春梅. Nrf2信号通路作为虾青素治疗新靶点的研究进展[J]. 国际医药卫生导报,2024,30(1): 20-24.DOI: 10.3760/cma.j.issn.1007-1245.2024.01.004. [40] LvB, XingS, WangZ, et al. NRF2 inhibitors: recent progress, future design and therapeutic potential[J]. Eur J Med Chem, 2024, 279: 116822.DOI: 10.1016/j.ejmech.2024.116822. -

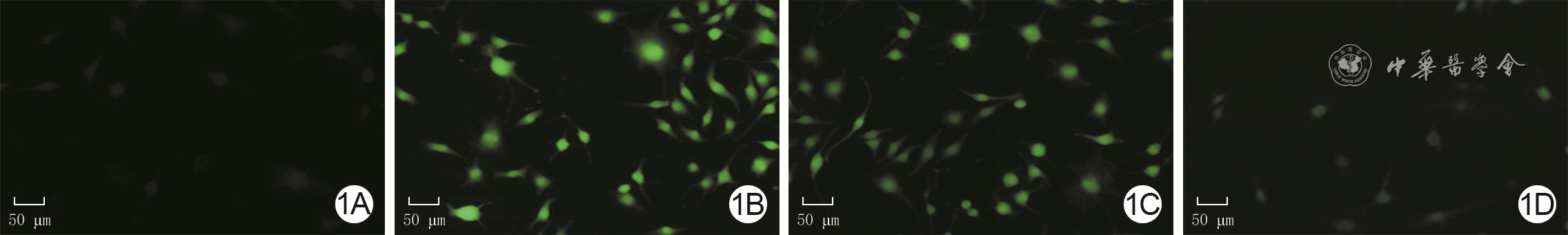
图 1 4组人成纤维细胞培养48 h后的活性氧水平 2',7'-二氯二氢荧光素二乙酸酯×100。1A、1B、1C、1D.分别为对照组、高糖组、低虾青素组、高虾青素组,图1B活性氧水平明显高于图1A、1C、1D,图1C活性氧水平明显高于图1D
注:对照组细胞行常规培养,高糖组细胞用终物质的量浓度30 mmol/L葡萄糖处理,低虾青素组、高虾青素组细胞分别用终物质的量浓度25、50 μmol/L虾青素预处理后再同高糖组处理;绿色荧光表示活性氧

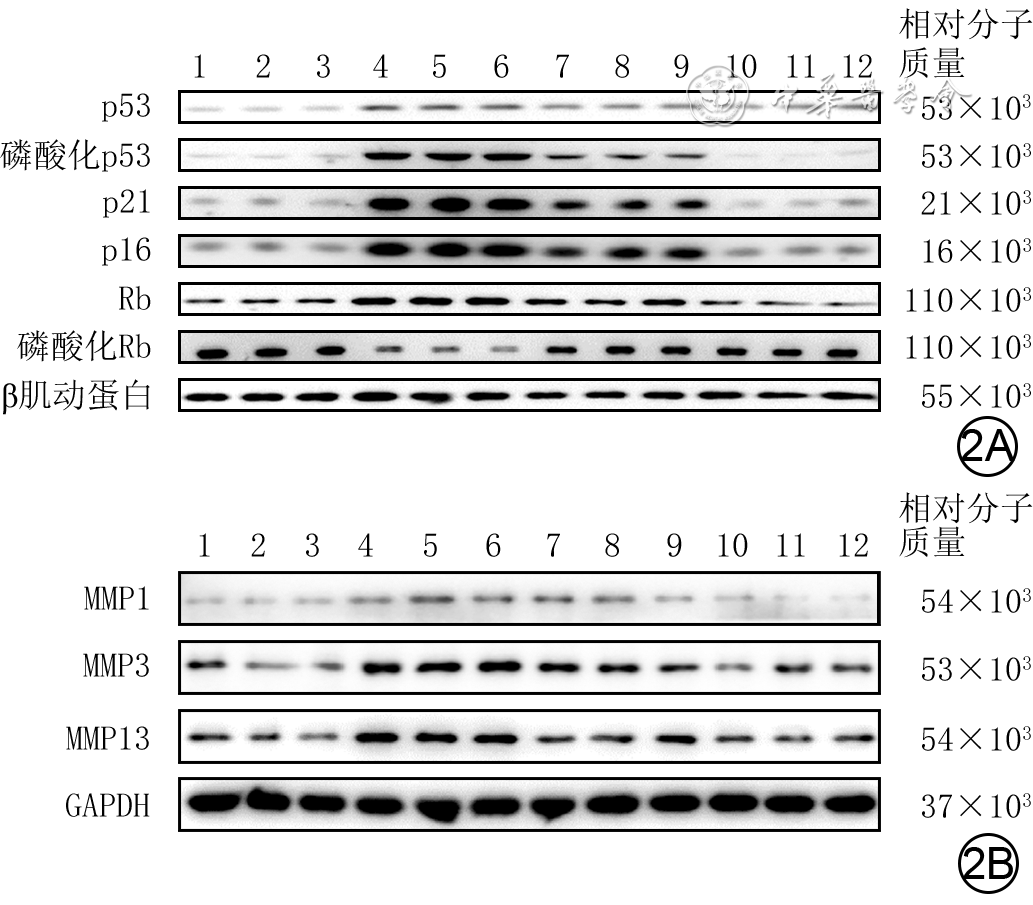
图 2 蛋白质印迹法检测的4组人成纤维细胞培养48 h后部分衰老相关蛋白表达水平。2A.细胞中p53、磷酸化p53、p21、p16、Rb、磷酸化Rb;2B.细胞中MMP1、MMP3、MMP13
注:条带上方1~3、4~6、7~9、10~12分别指示对照组、高糖组、低虾青素组、高虾青素组;对照组细胞行常规培养,高糖组细胞用终物质的量浓度30 mmol/L葡萄糖处理,低虾青素组、高虾青素组细胞分别用终物质的量浓度25、50 μmol/L虾青素预处理后再同高糖组处理;MMP为基质金属蛋白酶,GAPDH为3-磷酸甘油醛脱氢酶

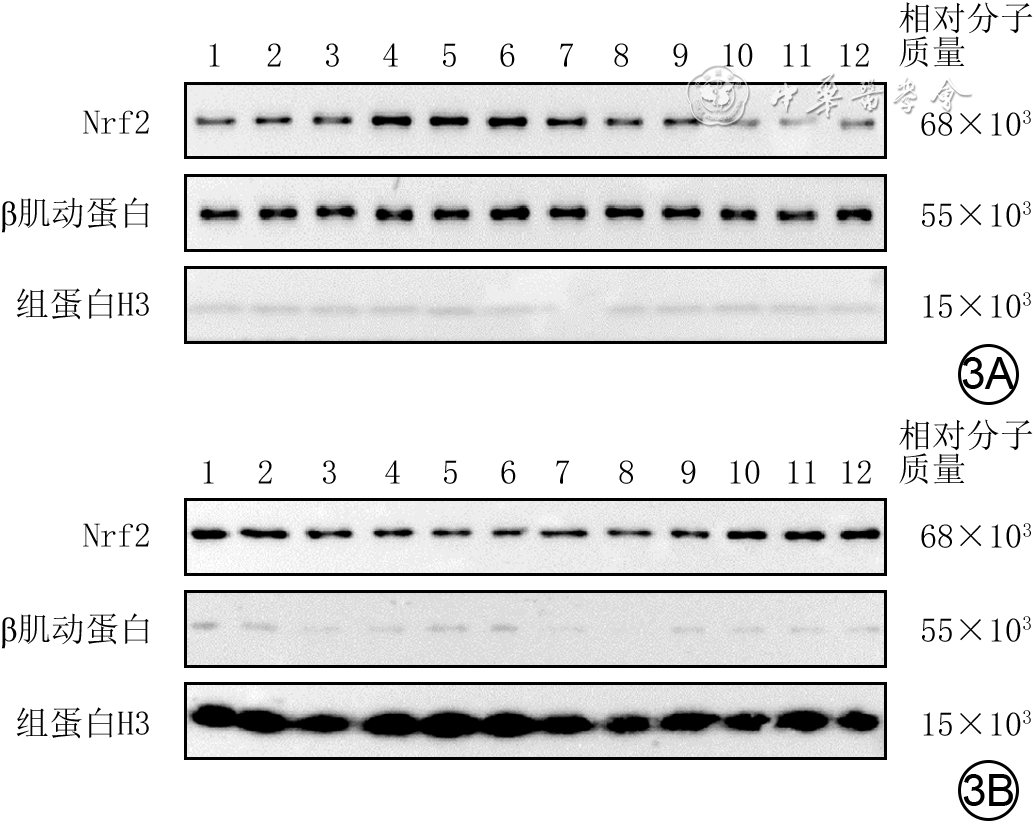
图 3 蛋白质印迹法检测的4组人成纤维细胞培养48 h后细胞质和细胞核中Nrf2蛋白表达水平。3A.细胞质;3B.细胞核
注:Nrf2为核转录因子红系2相关因子2;条带上方1~3、4~6、7~9、10~12分别指示对照组、高糖组、低虾青素组、高虾青素组;对照组细胞行常规培养,高糖组细胞用终物质的量浓度30 mmol/L葡萄糖处理,低虾青素组、高虾青素组细胞分别用终物质的量浓度25、50 μmol/L虾青素预处理后再同高糖组处理

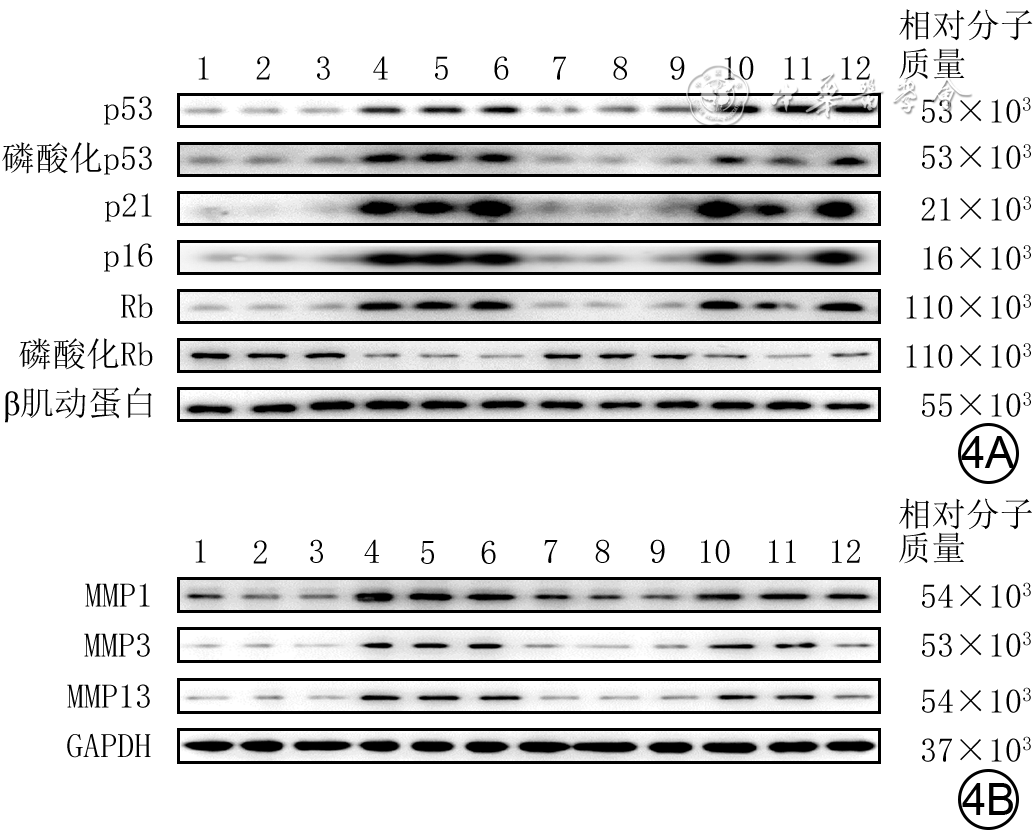
图 4 蛋白质印迹法检测的4组人成纤维细胞培养48 h后部分衰老相关蛋白表达水平。4A.细胞中p53、磷酸化p53、p21、p16、Rb、磷酸化Rb;4B.细胞中MMP1、MMP3、MMP13
注:条带上方1~3、4~6、7~9、10~12分别指示对照组、高糖组、高虾青素组、高虾青素+ML385组;对照组细胞行常规培养,高糖组细胞用终物质的量浓度30 mmol/L葡萄糖处理,高虾青素组和高虾青素+ML385组细胞分别用终物质的量浓度50 μmol/L虾青素、终物质的量浓度50 μmol/L虾青素+10 μmol/L ML385预处理后再同高糖组处理;MMP为基质金属蛋白酶,GAPDH为3-磷酸甘油醛脱氢酶

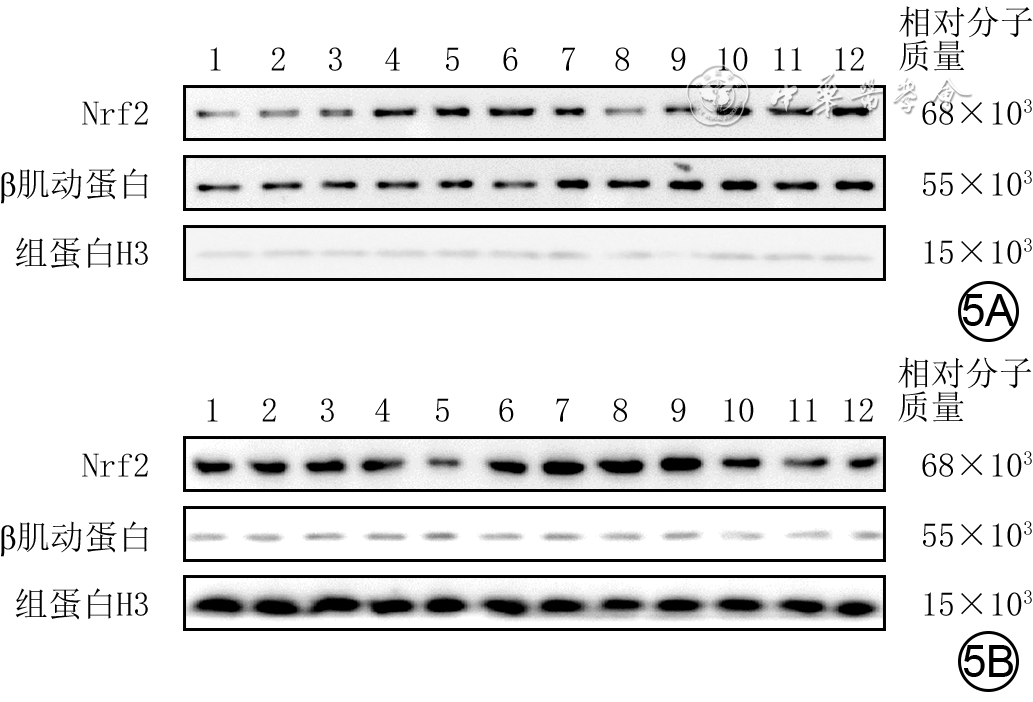
图 5 蛋白质印迹法检测的4组人成纤维细胞培养48 h后细胞质和细胞核中Nrf2蛋白表达水平。5A.细胞质;5B.细胞核
注:Nrf2为核转录因子红系2相关因子2;条带上方1~3、4~6、7~9、10~12分别指示对照组、高糖组、高虾青素组、高虾青素+ML385组;对照组细胞行常规培养,高糖组细胞用终物质的量浓度30 mmol/L葡萄糖处理,高虾青素组和高虾青素+ML385组细胞分别用终物质的量浓度50 μmol/L虾青素、终物质的量浓度50 μmol/L虾青素+10 μmol/L ML385预处理后再同高糖组处理
Table 1. 4组人成纤维细胞培养48 h后氧化损伤相关指标水平比较(
组别 样本数 活性氧 丙二醛 谷胱甘肽 SOD CAT GSH-Px 对照组 3 3.3±0.6 4.0±0.6 22.0±3.1 30.3±4.6 37.0±1.3 181±12 高糖组 3 29.6±1.0 11.4±1.1 7.8±2.3 11.3±1.0 15.4±3.3 74±4 低虾青素组 3 20.8±1.4 7.7±0.6 14.4±0.6 17.5±1.1 23.4±2.4 108±9 高虾青素组 3 7.6±0.7 4.7±0.4 21.7±3.0 24.4±2.7 36.4±0.8 173±8 F值 473.30 62.48 22.74 27.39 69.06 105.40 P 值 <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 P1值 <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 P2值 <0.001 0.002 0.066 0.144 0.013 0.008 P3值 <0.001 <0.001 <0.001 0.002 <0.001 <0.001 P4值 <0.001 0.007 0.040 0.091 <0.001 <0.001 注:对照组细胞行常规培养,高糖组细胞用终物质的量浓度30 mmol/L葡萄糖处理,低虾青素组、高虾青素组细胞分别用终物质的量浓度25、50 μmol/L虾青素预处理后再同高糖组处理;SOD为超氧化物歧化酶,CAT为过氧化氢酶,GSH-Px为谷胱甘肽过氧化物酶;F值、P值为组间各指标总体比较所得;P1值、P2值、P3值、P4值分别为对照组与高糖组、高糖组与低虾青素组、高糖组与高虾青素组、低虾青素组与高虾青素组各指标比较所得  下载: 导出CSV
下载: 导出CSV
Table 2. 4组人成纤维细胞培养48 h后部分衰老相关蛋白表达水平比较(
组别 样本数 p53 p21 磷酸化p53 p16 Rb 磷酸化Rb MMP1 MMP3 MMP13 对照组 3 1.00±0.05 1.00±0.08 1.00±0.26 1.00±0.12 1.000±0.127 1.00±0.12 1.00±0.19 1.00±0.29 1.00±0.13 高糖组 3 4.39±0.17 5.30±0.15 6.95±0.11 3.71±0.36 1.915±0.186 0.36±0.05 2.71±0.45 2.37±0.13 2.20±0.23 低虾青素组 3 3.29±0.24 3.53±0.10 3.69±0.23 3.61±0.30 1.838±0.154 0.96±0.04 2.00±0.35 1.69±0.37 1.38±0.27 高虾青素组 3 3.35±0.57 1.01±0.45 0.94±0.14 1.05±0.17 0.868±0.190 0.87±0.05 0.88±0.38 1.13±0.33 1.08±0.11 F值 59.29 221.48 630.87 106.64 32.50 51.87 17.66 13.49 23.52 P值 <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 0.002 <0.001 P1值 <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 0.002 0.003 <0.001 P2值 0.018 <0.001 <0.001 >0.999 >0.999 <0.001 0.246 0.127 0.005 P3值 0.025 <0.001 <0.001 <0.001 <0.001 <0.001 0.001 0.005 <0.001 P4值 >0.999 <0.001 <0.001 <0.001 <0.001 0.870 0.030 0.305 0.601 注:对照组细胞行常规培养,高糖组细胞用终物质的量浓度30 mmol/L葡萄糖处理,低虾青素组、高虾青素组细胞分别用终物质的量浓度25、50 μmol/L的虾青素预处理后再同高糖组处理;F值、P值为组间各指标总体比较所得;P1值、P2值、P3值、P4值分别为对照组与高糖组、高糖组与低虾青素组、高糖组与高虾青素组、低虾青素组与高虾青素组各指标比较所得;MMP为基质金属蛋白酶
下载: 导出CSV
Table 3. 4组人成纤维细胞培养48 h后细胞质和细胞核中Nrf2蛋白表达水平比较(
组别 样本数 细胞质中Nrf2 细胞核中Nrf2 对照组 3 1.000±0.147 1.000±0.018 高糖组 3 1.717±0.144 0.585±0.097 低虾青素组 3 0.906±0.206 0.795±0.056 高虾青素组 3 0.397±0.077 1.139±0.141 F值 39.04 21.59 P值 <0.001 <0.001 P1值 0.002 0.003 P2值 0.001 0.130 P3值 <0.001 <0.001 P4值 0.020 0.010 注:对照组细胞行常规培养,高糖组细胞用终物质的量浓度30 mmol/L葡萄糖处理,低虾青素组、高虾青素组细胞分别用终物质的量浓度25、50 μmol/L虾青素预处理后再同高糖组处理;Nrf2为核转录因子红系2相关因子2;F值、P值为组间各指标总体比较所得;P1值、P2值、P3值、P4值分别为对照组与高糖组、高糖组与低虾青素组、高糖组与高虾青素组、低虾青素组与高虾青素组各指标比较所得
下载: 导出CSV
Table 4. 4组人成纤维细胞培养48 h后部分衰老相关蛋白表达水平比较(
组别 样本数 p53 p21 磷酸化p53 p16 Rb 磷酸化Rb MMP1 MMP3 MMP13 对照组 3 1.00±0.07 1.00±0.21 1.00±0.03 1.00±0.26 1.000±0.061 1.00±0.10 1.00±0.27 1.00±0.12 1.00±0.12 高糖组 3 3.31±0.36 7.30±1.04 2.46±0.12 2.97±0.13 4.175±0.070 0.38±0.04 5.24±0.30 5.14±0.12 4.53±0.29 高虾青素组 3 2.16±0.56 2.60±0.10 0.90±0.09 1.29±0.27 1.036±0.008 0.91±0.09 1.54±0.22 1.55±0.37 1.24±0.34 高虾青素+ML385组 3 4.97±0.29 6.54±0.77 2.15±0.40 2.27±0.22 3.371±0.489 0.40±0.10 4.10±0.43 3.82±1.56 2.75±0.81 F值 63.55 64.54 41.77 48.19 128.00 42.20 122.50 17.26 36.44 P值 <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 P1值 <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 0.001 <0.001 P2值 0.031 <0.001 <0.001 <0.001 <0.001 <0.001 <0.001 0.004 <0.001 P3值 <0.001 <0.001 <0.001 0.004 <0.001 <0.001 <0.001 0.053 0.025 注:对照组细胞行常规培养,高糖组细胞用终物质的量浓度30 mmol/L葡萄糖处理,高虾青素组和高虾青素+ML385组细胞分别用终物质的量浓度50 μmol/L虾青素、终物质的量浓度50 μmol/L虾青素+10 μmol/L ML385预处理后再同高糖组处理;F值、P值为组间各指标总体比较所得;P1值、P2值、P3值分别为对照组与高糖组、高糖组与高虾青素组、高虾青素组与高虾青素+ML385组各指标比较所得;MMP为基质金属蛋白酶
下载: 导出CSV
Table 5. 4组人成纤维细胞培养48 h后细胞质和细胞核中Nrf2蛋白表达水平比较(
组别 样本数 细胞质中Nrf2 细胞核中Nrf2 对照组 3 1.000±0.172 1.000±0.030 高糖组 3 1.983±0.311 0.931±0.187 高虾青素组 3 0.917±0.275 1.494±0.129 高虾青素+ML385组 3 1.513±0.077 0.969±0.103 F值 14.19 13.34 P值 0.001 0.002 P1值 0.004 0.999 P2值 0.003 0.004 P3值 0.075 0.005 注:对照组细胞行常规培养,高糖组细胞用终物质的量浓度30 mmol/L葡萄糖处理,高虾青素组和高虾青素+ML385组细胞分别用终物质的量浓度50 μmol/L虾青素、终物质的量浓度50 μmol/L虾青素+10 μmol/L ML385预处理后再同高糖组处理;Nrf2为核转录因子红系2相关因子2;F值、P值为组间各指标总体比较所得;P1值、P2值、P3值分别为对照组与高糖组、高糖组与高虾青素组、高虾青素组与高虾青素+ML385组各指标比较所得
下载: 导出CSV
-

 下载:
下载:


计量
- 文章访问数: 838
- HTML全文浏览量: 291
- PDF下载量: 29
- 被引次数: 0
