Liu XX,Liu DW.Research advances on the effects of RNA N6-methyladenosine modification in the relevant pathophysiological processes of wound repair[J].Chin J Burns Wounds,2022,38(10):989-993.DOI: 10.3760/cma.j.cn501120-20210804-00267.
Citation:
Yu XP,Liu YK,Ma X,et al.Effect of deep dermal tissue dislocation injury on skin fibrosis in pig[J].Chin J Burns Wounds,2022,38(11):1057-1065.DOI: 10.3760/cma.j.cn501120-20210831-00301.
Liu XX,Liu DW.Research advances on the effects of RNA N6-methyladenosine modification in the relevant pathophysiological processes of wound repair[J].Chin J Burns Wounds,2022,38(10):989-993.DOI: 10.3760/cma.j.cn501120-20210804-00267.
Citation:
Yu XP,Liu YK,Ma X,et al.Effect of deep dermal tissue dislocation injury on skin fibrosis in pig[J].Chin J Burns Wounds,2022,38(11):1057-1065.DOI: 10.3760/cma.j.cn501120-20210831-00301.
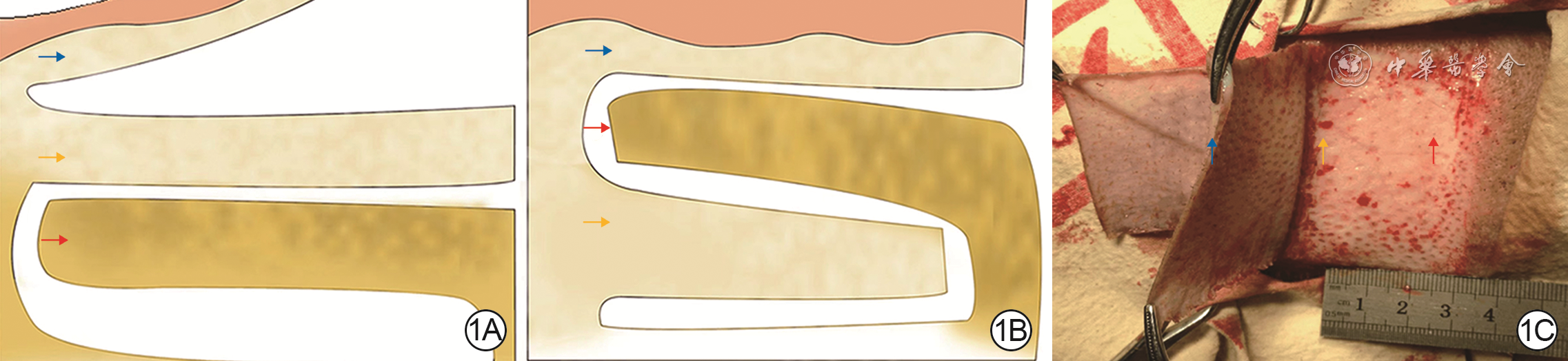
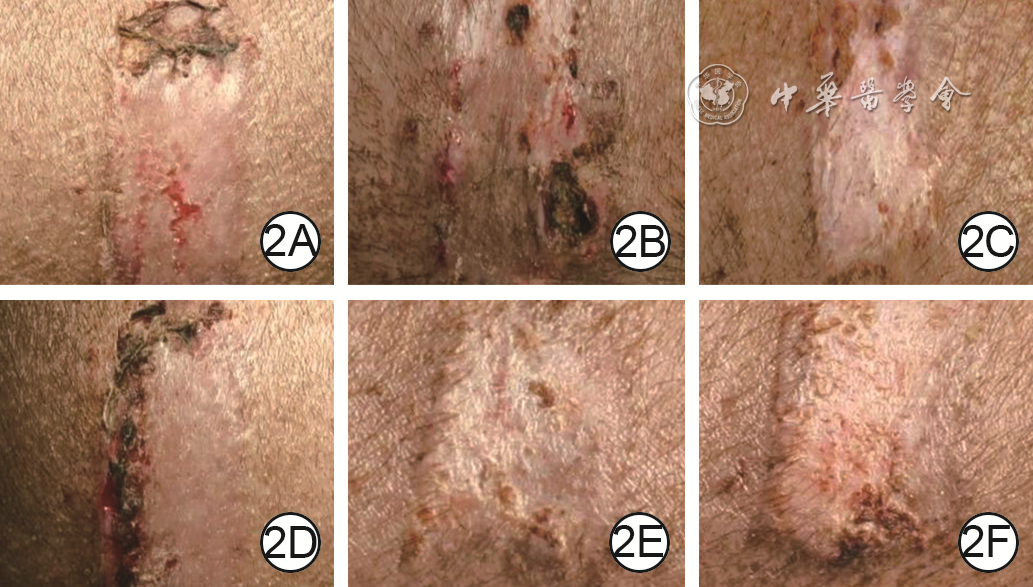
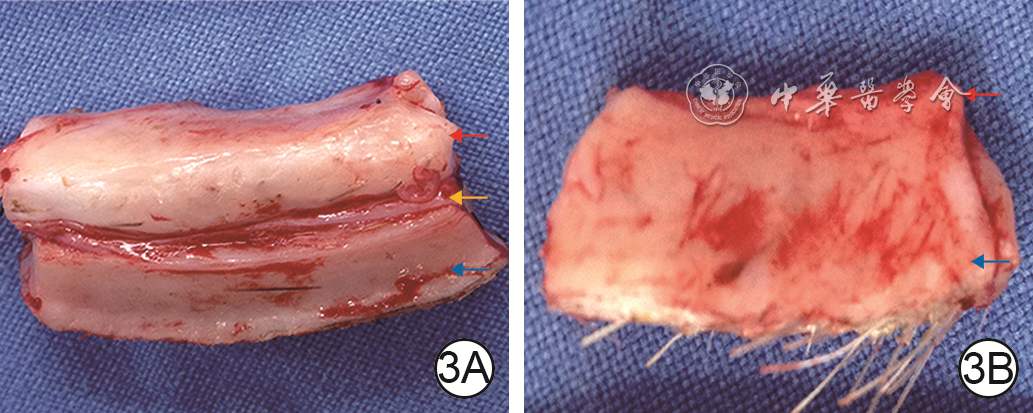
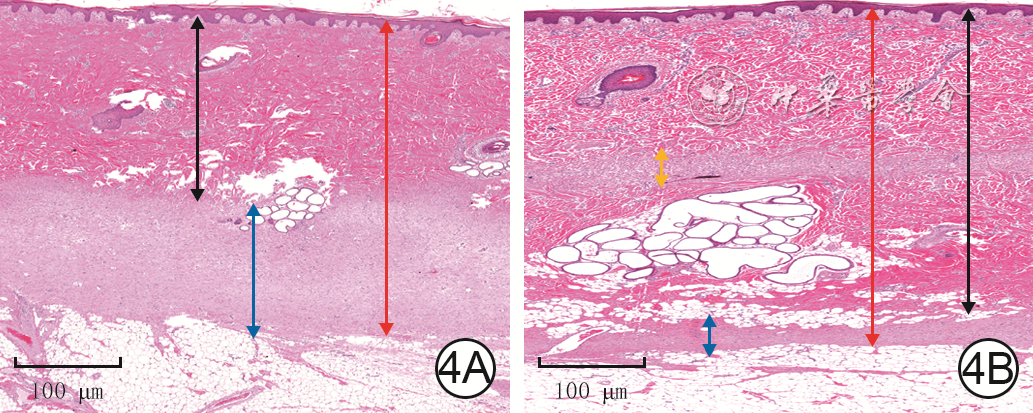
Objective To explore the effect of deep dermal tissue dislocation injury on skin fibrosis in pig, in order to provide some theoretical basis for burn scar treatment.Methods The experimental research method was applied. Six 2-month-old female Duroc pigs were taken. Fifteen operative areas on the right dorsum of pigs on which medium-thick skin grafts and deep dermal tissue slices were cut and re-implanted were included into dermal in situ reimplantation group, and fifteen operative areas on the left dorsum of pigs on which medium-thick skin grafts and deep dermal tissue slices were cut and the deep dermal tissue slice was placed under the fat layer were included into the dermal dislocation group. The hair growth in the operative areas on post-injury day (PID) 7, 14, and 21 and the cross-sectional structure on PID 14 were observed in the two groups. On PID 7, 14, and 21, the skin thickness (the distance from the epidermis to the upper edge of the fat), the dermal thickness (the distance from the lower edge of the epidermis to the upper edge of the fat, excluding the fibrotic tissue thickness between the dermis and the fat), and the fibrosis tissue thickness of the dermis-fat interface (from the lower edge of the deep dermis to the upper edge of the fat in dermal in situ reimplantation group and from the lower edge of the superficial dermis to the upper edge of the fat in dermal dislocation group) in the operative areas were measured and compared between the two groups; the fibrotic tissue thickness at the dermal cutting interface (from the lower edge of the superficial dermis to the upper edge of the deep dermis) in the operative areas in dermal in situ reimplantation group was measured and compared with the fibrotic tissue thickness at the dermal-fat interface. Sirius red staining was performed to observe and compare the type Ⅰ and Ⅲ collagen content in the dermal-fat interface in the operative areas between the 2 groups and between the dermal cutting interface and dermal-fat interface in the operative areas in dermal in situ reimplantation group. Immunohistochemical staining was performed to observe the positive expressions of proliferating cell nuclear antigen (PCNA), transforming growth factor β1 (TGF-β1), fibroblast growth factor 2 (FGF-2), and hepatocyte growth factor (HGF) in the operative areas in the two groups. The sample number was 6. Data were statistically analyzed with independent sample t test.Results On PID 7, 14, and 21, the hairs in the operative areas in dermal in situ reimplantation group were denser than those in dermal dislocation group. On PID 14, the skin cross section in the operative areas in dermal dislocation group showed a "sandwich"-like structure, while the skin cross section in the operative areas in dermal in situ reimplantation group had normal structure. On PID 7, 14, and 21, the skin thickness in the operative areas in dermal dislocation group was (4 234±186), (4 688±360), and (4 548±360) μm, respectively, which was close to (4 425±156), (4 714±141), and (4 310±473) μm in dermal in situ reimplantation group (P>0.05); the dermal thickness in the operative areas in dermal dislocation group was significantly thinner than that in dermal in situ reimplantation group (with t values of -9.73, -15.85, and -15.41, respectively, P<0.01); the fibrotic tissue thickness at the dermal-fat interface in the operative areas in dermal dislocation group was significantly thicker than that in dermal in situ reimplantation group (with t values of 14.48, 20.58, and 15.67, respectively, P<0.01); there was no statistically significant difference between the fibrotic tissue thickness at the dermal-fat interface and the dermal cutting interface in the operative areas in dermal in situ reimplantation group (P>0.05). On PID 7, 14, 21, the type Ⅲ collagen content in the dermal-fat interface in the operative areas in dermal dislocation group was increased significantly compared with that in dermal in situ replantation group (with t values of 2.65, 0.61, and 7.39, respectively, P<0.05 or P<0.01), whereas there were no statistically significant differences in the type Ⅰ collagen content at the dermal-fat interface in the operative areas between the 2 groups (P>0.05) and the type Ⅰ and Ⅲ collagen content between the dermal-fat interface and the dermal cutting interface in the operative areas in dermal in situ reimplantation group (P>0.05). On PID 7, 14, and 21, PCNA, TGF-β1, FGF-2, and HGF were positively expressed in the superficial dermis and adipose tissue in the operative areas in dermal dislocation group, while PCNA, TGF-β1, FGF-2, and HGF were positively expressed in the superficial dermis, deep dermis, and adipose tissue in the operative areas in dermal in situ reimplantation group.Conclusions Inadequate intrinsic thickness of dermal tissue is the key factor causing fibrosis, and the biological purpose of fibrosis is to "compensate" the intrinsic thickness of the skin. Besides, adipose tissue may also be an important component of fibrotic skin repair.
DingJ,MaZ,ShankowskyHA,et al.Deep dermal fibroblast profibrotic characteristics are enhanced by bone marrow-derived mesenchymal stem cells[J].Wound Repair Regen,2013,21(3):448-455.DOI: 10.1111/wrr.12046.
[3]
HonardoustD,DingJ,VarkeyM,et al.Deep dermal fibroblasts refractory to migration and decorin-induced apoptosis contribute to hypertrophic scarring[J].J Burn Care Res,2012,33(5):668-677.DOI: 10.1097/BCR.0b013e31824088e3.
[4]
Ali-BaharM,BauerB,TredgetEE,et al.Dermal fibroblasts from different layers of human skin are heterogeneous in expression of collagenase and types I and III procollagen mRNA[J].Wound Repair Regen,2004,12(2):175-182.DOI: 10.1111/j.1067-1927.2004.012110.x.
[5]
WangJ,DoddC,ShankowskyHA,et al.Deep dermal fibroblasts contribute to hypertrophic scarring[J].Lab Invest,2008,88(12):1278-1290.DOI: 10.1038/labinvest.2008.101.
HonardoustD,VarkeyM,MarcouxY,et al.Reduced decorin, fibromodulin, and transforming growth factor-β3 in deep dermis leads to hypertrophic scarring[J].J Burn Care Res,2012,33(2):218-227.DOI: 10.1097/BCR.0b013e3182335980.
[11]
KimWS,ParkBS,SungJH,et al.Wound healing effect of adipose-derived stem cells: a critical role of secretory factors on human dermal fibroblasts[J].J Dermatol Sci,2007,48(1):15-24.DOI: 10.1016/j.jdermsci.2007.05.018.
[12]
ShiHX,LinC,LinBB,et al.The anti-scar effects of basic fibroblast growth factor on the wound repair in vitro and in vivo[J].PLoS One,2013,8(4):e59966.DOI: 10.1371/journal.pone.0059966.
[13]
MarangoniRG,KormanBD,WeiJ,et al.Myofibroblasts in murine cutaneous fibrosis originate from adiponectin-positive intradermal progenitors[J].Arthritis Rheumatol,2015,67(4):1062-1073.DOI: 10.1002/art.38990.
[14]
PanX,ChenZ,HuangR,et al.Transforming growth factor β1 induces the expression of collagen type I by DNA methylation in cardiac fibroblasts[J].PLoS One,2013,8(4):e60335.DOI: 10.1371/journal.pone.0060335.
[15]
WikanN,PotikanondS,NimlamoolW.Alpinetin suppresses effects of TGF-β1 on stimulating the production and organization of fibrotic markers in human primary dermal fibroblasts[J].Cells,2022,11(17):2731.DOI: 10.3390/cells11172731.
[16]
KimWS,ParkBS,ParkSH,et al.Antiwrinkle effect of adipose-derived stem cell: activation of dermal fibroblast by secretory factors[J].J Dermatol Sci,2009,53(2):96-102.DOI: 10.1016/j.jdermsci.2008.08.007.
[17]
van den BogaerdtAJ,van der VeenVC,van ZuijlenPP,et al.Collagen cross-linking by adipose-derived mesenchymal stromal cells and scar-derived mesenchymal cells: are mesenchymal stromal cells involved in scar formation?[J].Wound Repair Regen,2009,17(4):548-558.DOI: 10.1111/j.1524-475X.2009.00501.x.
[18]
TredgetEE.Pathophysiology and treatment of fibroproliferative disorders following thermal injury[J].Ann N Y Acad Sci,1999,888:165-182.DOI: 10.1111/j.1749-6632.1999.tb07955.x.
[19]
XueM,ZhaoR,MarchL,et al.Dermal fibroblast heterogeneity and its contribution to the skin repair and regeneration[J].Adv Wound Care (New Rochelle),2022,11(2):87-107.DOI: 10.1089/wound.2020.1287.
[20]
WeiJJ,TangL,ChenLL,et al.Mesenchymal stem cells attenuates TGF-β1-induced EMT by increasing HGF expression in HK-2 cells[J].Iran J Public Health,2021,50(5):908-918.DOI: 10.18502/ijph.v50i5.6108.
[21]
WuZY,ZhangHJ,ZhouZH,et al.The effect of inhibiting exosomes derived from adipose-derived stem cells via the TGF-β1/Smad pathway on the fibrosis of keloid fibroblasts[J].Gland Surg,2021,10(3):1046-1056.DOI: 10.21037/gs-21-4.
ChengTL,ChenPK,HuangWK,et al.Plasminogen/thrombomodulin signaling enhances VEGF expression to promote cutaneous wound healing[J].J Mol Med (Berl),2018,96(12):1333-1344.DOI: 10.1007/s00109-018-1702-1.
[25]
StanojcicM,AbdullahiA,RehouS,et al.Pathophysiological response to burn injury in adults[J].Ann Surg,2018,267(3):576-584.DOI: 10.1097/SLA.0000000000002097.
WangJ,CaiJ,ZhangQ,et al.Fat transplantation induces dermal adipose regeneration and reverses skin fibrosis through dedifferentiation and redifferentiation of adipocytes[J].Stem Cell Res Ther,2022,13(1):499.DOI: 10.1186/s13287-022-03127-0.
Liu XX,Liu DW.Research advances on the effects of RNA N6-methyladenosine modification in the relevant pathophysiological processes of wound repair[J].Chin J Burns Wounds,2022,38(10):989-993.DOI: 10.3760/cma.j.cn501120-20210804-00267.
Liu XX,Liu DW.Research advances on the effects of RNA N6-methyladenosine modification in the relevant pathophysiological processes of wound repair[J].Chin J Burns Wounds,2022,38(10):989-993.DOI: 10.3760/cma.j.cn501120-20210804-00267.





 下载:
下载:





 DownLoad:
DownLoad: